

家禽和其他单胃动物中的纳米硒同化和作用:肠道微生物群是答案吗?
摘要
最近,一篇关于纳米硒在牲畜和鱼类营养中的作用的综合评论论文发表在纳米级研究快报上。作者详细描述了与纳米硒生产相关的问题及其在动物工业和医学中的可能应用。然而,纳米硒作用的分子机制没有被描述,纳米硒如何转化为活性硒蛋白的问题也没有解决。肠道微生物群似乎可以通过以下硒蛋白的合成将纳米硒转化为亚硒酸盐、H2Se 或硒磷酸盐。这种可能性需要进一步详细研究,纳米硒作为动物/家禽/鱼类营养中硒的来源的优缺点有待批判性评估。
背景
最近,一篇关于纳米硒在牲畜和鱼类营养中的作用的综合评论论文发表在纳米级研究快报 [1] 上。作者非常详细地描述了与纳米硒生产相关的问题及其在动物工业和医学中的可能应用。事实上,众所周知,由于这些粒子的新特性,许多以纳米粒子形式呈现的分子具有不寻常的行为。事实上,在大多数与动物营养相关的已发表工作中,纳米粒子的尺寸小于 100 纳米。在这种状态下,粒子的超小尺寸使其能够穿透许多生物屏障,并用作各种元素的传递系统,包括银、钛、硒等。
在这方面,硒特别有趣,因为事实证明,全世界大部分地区都缺乏这种元素 [2]。在许多情况下,缺乏是人为造成的,因为土壤中硒的含量变化很大,更重要的是,土壤中硒的可用性变化更大。事实上,农业实践导致的土壤酸化以及含硫肥料的使用大大降低了植物对硒的可用性,并导致牲畜的硒消耗量低,其中谷物(小麦、玉米、大麦)和油籽(大豆)构成饮食的主要部分。因此,在家禽、农场动物和鱼类的商业饮食中补充硒成为一种普遍做法,并自 1970 年代开始使用。事实上,维生素-矿物质预混料是用于鸡蛋、肉/鱼和牛奶商业生产的硒的主要来源。
与纳米硒故事相关的是补充剂中硒的来源。事实上,硒的主要补充形式是亚硒酸钠,它是铜生产的副产品。经过 40 多年的使用,亚硒酸钠已经清楚地展示了其优点和缺点。事实上,由于商业饲料硒的补充,具有缺乏临床症状的牲畜的硒缺乏症几乎消失了。反刍动物例外,因为在酸性瘤胃环境中亚硒酸钠会沉淀,而且由于饲料添加剂使用的技术限制;世界范围内仍然存在硒缺乏症,并且使用硒注射和大丸剂等应用。然而,正如上述评论中正确提到的那样,硒在商业相关的压力条件下极其重要,特别是对于高产农场动物和家禽的现代遗传学。
今天,畜牧业正在从预防营养缺乏转向满足动物对重要营养物质(包括硒)的确切需求。事实上,“精确营养”是描述这一概念的术语。在这种情况下,很明显亚硒酸钠(以及硒酸盐)作为饲料补充剂具有一系列缺点。首先,它是一种反应性很强的化合物,可以在饲料/预混料中被各种营养物质(包括抗坏血酸和一些饲料成分)还原成不可利用的金属形式。它也可以溶解在饲料水分中并转化为挥发性化合物而损失掉。其次,亚硒酸钠以剂量依赖性方式具有促氧化特性,这可能对动物/小鸡肠道产生负面影响。最后,亚硒酸钠形式的硒很难通过胎盘转移到鸡蛋中,从而无法在体内建立硒储备,当硒需求增加但饲料消耗通常会减少时,硒可用于应激条件下。亚硒酸钠过量对动物和人类也是有毒的。然而,我们不同意上述评论 [1] 中的结论,即硒毒性是动物工业的一个问题。它仅与计算或混合饲料中的人为错误有关。硒的安全范围非常窄(家禽和猪的硒补充剂量通常为 0.3 ppm,而亚硒酸钠剂量超过 1-2 ppm 时会观察到负面影响),但现代饲料厂设备提供了良好的饲料混合机会,避免毒性问题。
分析科学的进步推动了我们发现主要饲料成分中硒的主要形式是硒化硒,占玉米、大豆、小麦、大麦等硒总量的 50% 以上 [2]。因此,在进化过程中,动物的消化系统适应了这种形式的硒,因此,SeMet 可以更有效地从饮食中吸收,并且非特异性地掺入身体蛋白质中以建立硒储备。事实上,市场上出现了一系列有机硒源,包括 Se-Yeast、纯 SeMet 和 OH-SeMet(2-羟基-4-(甲硫基)丁酸-Se),其中 OH-SeMet 显示出最大的功效。
纳米硒问题的主要知识缺乏与其代谢有关,特别是其转化为 H2Se,随后 SeCys 合成并结合到硒蛋白中。在综述 [1] 中,只提到了极少数硒蛋白,包括谷胱甘肽过氧化物酶,而在人类和动物中至少发现了 25 种硒蛋白。人们普遍认为,硒在人类/动物营养中的主要作用与具有独特催化特性的硒蛋白的合成有关,其中一半以上参与维持氧化还原平衡和抗氧化防御 [3]。在上述评论 [1] 中,纳米硒的直接抗氧化特性(减少 ROS)被认为是其作用的可能机制。然而,包括肝脏和肌肉在内的主要动物/家禽组织中的硒浓度通常不超过 800-900 ng/g 新鲜组织,在 10 μM 的范围内,而在血浆中,硒水平约为 0.2-0.3 μg/ ml 或 2-3 μM,而在被引用最多的致力于纳米硒体外抗氧化特性的工作中,测试了硒浓度并显示出高 5-10 倍的抗氧化效果 [4]。此外,要使抗氧化剂化合物成为有效的自由基清除剂,重要的是要在正确的时间、正确的地点使用正确浓度的抗氧化剂。这会使问题进一步复杂化,因此,纳米硒似乎不太可能在生物系统中产生直接的 AO 效应。
因此,类似于动物饮食中使用的其他形式的硒,纳米硒的抗氧化作用与硒蛋白基因表达和蛋白质合成有关。事实上,在评论 [1] 中,有几篇参考文献证实了纳米硒对 GSH-Px 活性的积极影响。现在的问题是如何 纳米硒转化为活性硒蛋白。在综述 [1] 中,有一个建议(没有参考),纳米硒可以通过以下硒蛋白合成转化为硒代磷酸盐。这个建议应该得到实验证明。纳米硒可以转化为SeMet的第二个建议从根本上是错误的,因为SeMet不能在人/动物体内合成;只有植物和细菌才能产生它[5]。
有一些非常有吸引力的建议,即肠道微生物群可以将纳米硒氧化成亚硒酸盐/硒酸盐或通过以下硒蛋白的合成将其还原成 H2Se [6, 7]。最近,已经提供了一些实验证据来证明纳米硒颗粒在被吸收之前在微生物群存在的情况下可以在肠道中溶解并氧化成硒的无机含氧阴离子[6]。此外,还提出了一个假设示意图,显示了内源性硒纳米颗粒 (SeNPs) 的细胞内动态循环 [7]。事实上,有人提出元素硒可以被超氧自由基再氧化成亚硒酸盐。肠道微生物群也有可能执行/加速这一过程。事实上,Se 的四种主要生物转化已被证明在自然界发生,包括还原、氧化、甲基化和去甲基化 [8]。过去三年积累的数据清楚地表明,微生物通过参与氧化和还原反应,在环境中的硒循环中发挥着重要作用 [9]。有趣的是,Se 0 的微生物氧化 到Se 4+ 由一群身份不明的自养细菌在 90 多年前被发现 [10]。此外,异养细菌巨大芽孢杆菌将元素硒氧化为亚硒酸盐 ,从土壤中分离出来的报道要晚得多[11]。确实,Se 0 土壤中的氧化被证明以相对缓慢的速度发生,并且在自然界中主要是生物的,并且产生 SeO3 2− 和 SeO4 2− [12]。此外,元素硒(Se 0 ) 由化学异养菌和化学自养硫杆菌通过 75 确认 Se 0 作为示踪剂 [13]。作者表明,土壤泥浆能够氧化 Se 0 含 SeO3 2− 和 SeO4 2− 形成。有趣的是,通过高压灭菌或化学处理使土壤中的微生物失活抑制了该过程。此外,硫氧化细菌硫杆菌的培养 研究表明,ASN-1 可以使用酶对 Se(0) 进行氧化,这些酶用于从还原的硫化合物中产生能量 [13]。此外,可以氧化 Mn(II) 或 Fe(II) 的异养细菌 (Leptothrix MnB1) 被证明可以氧化 Se 0 随着 SeO3 2− 的形成 作为反应的主要产物。有趣的是,该反应显示依赖于电子供体,例如醋酸盐或葡萄糖 [13]。 Se 0 的氧化 由各种细菌引起的问题尚未得到充分解决,这是一个具有重要研究机会的领域。事实上,肠道中的多种微生物为各种硒转化提供了必要条件。例如,当纳米硒颗粒与乳酸菌(德氏乳杆菌 subsp。保加利亚 LB-12),产生有机硒化合物(主要是 SeCys 和 SeMet),并且纳米硒颗粒也被部分溶解和非代谢转化为无机硒,可能是在细菌细胞壁分泌的物质的帮助下 [14]。
或者,可以通过亚硒酸盐呼吸细菌(例如,硒化芽孢杆菌)将元素硒还原/转化为硒化物 )。上述细菌在电子供体乳酸不完全氧化为乙酸的情况下进行的反应如下[7]:
$$ {\mathrm{C}}_2{\mathrm{H}}_4{\mathrm{OHCOO}}^{-}+2{\mathrm{Se}}^0+2{\mathrm{H}}_2 \mathrm{O}\to {\mathrm{C}\mathrm{H}}_3{\mathrm{C}\mathrm{OO}}^{-}+2{\mathrm{H}\mathrm{Se}} ^{-}+{{\mathrm{H}\mathrm{CO}}_3}^{-}+3{\mathrm{H}}^{+} $$
因此,反应的自由能 (∆G
I
) 显示为 − 2.8 kcal/mol e
−
.这表明在硒化芽孢杆菌 ,还原机制涉及通过使用硒特异性异化酶进行能量保存 [15]。事实上,一种亚硒酸盐呼吸细菌,硒化芽孢杆菌 , 可以产生显着水平的 Se
− 2
(作为 HSe- 水溶液)使用 Se
0
作为底物[16]。较早前已证明专性嗜酸菌氧化亚铁硫杆菌可还原红硒 在酸性(pH 3)、厌氧条件下,H2Se 的产量为 0.03 μmol/mg 蛋白质/h [17]。此外,另一种厌氧菌Veillonella atypica 被证明能够还原硒氧阴离子以形成元素硒,细菌将其进一步还原以形成活性硒化物 [18]。有趣的是,在 45 多年前,有人描述了乳微球菌(Veillonella alcalescens ) 能够将胶体硒定量还原为 H2Se [19]。最近,编码 YedE 和 YedF 蛋白的基因被认为是参与包括细菌在内的原核生物中硒代谢的新候选基因 [20]。事实上,预测的硒转运蛋白 YedE 和氧化还原蛋白 YedF 都可能参与细菌细胞中硒的代谢转化。因此,证明了各种微生物氧化或还原元素硒的主要能力(表 1);然而,还需要进一步调查来回答此类反应是否发生在动物肠道中的问题。有趣的是,从表 1 中提到的所有细菌物种中,韦永氏菌 属的厌氧革兰氏阴性细菌 值得特别关注。 事实上,Veillonellae 存在于温血动物的消化道中 [21]。事实上,在食用动物中,Veillonella 作为胃肠道所有部分的土著居民([22] 和参考文献),包括鸡的上胃肠道 [23] 和盲肠 [24],定期检测到菌株。事实上,还需要进一步的研究来阐明纳米硒在动物胃肠道中可能转化的命运和机制。纳米硒代谢取决于纳米颗粒组成,包括纳米颗粒涂层剂。纳米硒参与硒蛋白合成的拟议方案如图 1 所示,对纳米硒代谢(包括吸收、分布和清除)的基本了解在动物/家禽科学中具有重要意义 [25]。 <图片>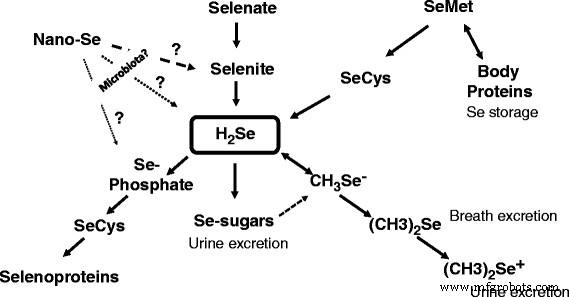
示意图模型显示了动物中各种形式的硒的代谢转化。表明肠道微生物群可以将纳米硒转化为亚硒酸盐、H2Se 或硒磷酸盐,并通过以下 SeCys 合成并掺入硒蛋白
哺乳动物/鸟类胃肠道含有数万亿的共生微生物,统称为微生物群 [26]。例如,鸡肠道由十二指肠、空肠、回肠、盲肠和结肠组成,上述肠道切片之间的微生物群浓度和组成存在显着差异[27]。有趣的是,盲肠的特征是由 Firmicutes 门主导的最复杂的微生物群落 , 拟杆菌 , 放线菌 和 Proteobacteria [28]。另一方面,在属水平上,所有肠道切片的主要微生物属均显示为乳酸菌 , 肠球菌 , 拟杆菌 , 和 棒状杆菌 [27]。此外,拟杆菌 被证明是盲肠中的优势组,而乳酸菌 主要存在于小肠部分(十二指肠、空肠和回肠;[27])。在这个复杂的肠道生态系统中,可能存在一系列微生物能够促进纳米硒颗粒的氧化/还原,这一假设有待进一步研究。
一方面,一些微生物会根据自身需要利用硒来合成微生物硒蛋白,并与宿主竞争这种元素。众所周知,硒是几乎所有细菌门中多种生物的重要元素;然而,似乎只有三分之一的特征细菌在其新陈代谢中使用这种元素 [29]。事实上,在硒缺乏的情况下,细菌和宿主之间存在着对可用硒的竞争,而无菌动物的硒需求量低于传统定植动物 [30]。最近的结果表明,膳食硒补充剂会影响现有微生物群的组成和多样性以及胃肠道微生物群落的建立 [31]。例如,与对照动物相比,饲喂富含硒的苜蓿干草的断奶小牛的鼻腔微生物群更丰富 [32]。各种形式和浓度的硒对肠道微生物群的可能影响有待进一步研究。
另一方面,微生物群似乎很可能主动吸收硒,这个过程取决于所使用的硒形式。事实上,与硒酸盐的摄取相比,结肠微生物群对 SeMet 的摄取显示出更有效 [33]。然而,尚未研究细菌组成和浓度是否会影响肠道对硒化合物的吸收。人们普遍认为,肠道微生物群负责通过甲基化和元素硒的形成来排出过量的硒 [34]。有趣的是,在喂食 SeMet 的大鼠中,在肠道的所有部分都发现了这种形式的硒。然而,益生菌处理后大鼠回肠、盲肠和结肠的硒含量显着高于相应切片[34]。这可能意味着唾液链球菌 , 鼠李糖乳杆菌 , 嗜酸乳杆菌 和乳双歧杆菌 主要在远处肠道中通过益生菌影响的硒代谢被输送到肠道。事实上,应该确定不同细菌在硒转化中的作用,以了解肠道每个部分在硒代谢和同化中的作用。此外,液体从大肠上部回流回小肠可能是小肠吸收 H2Se 和其他形式的硒的原因。事实上,最近有研究表明,鸡的特征是反向蠕动收缩,可以将标记物从泄殖腔移动到胼胝[35]。
可以预期纳米硒直接参与硒蛋白合成,因为在细胞培养中纳米硒增加了硒蛋白(GSH-Px 和 TR 活性)。然而,最近对硒蛋白表达优先级的理解可能会使这个问题复杂化。事实上,许多硒蛋白受氧化应激调节。特别是,GSH-Px1、GSH-Px4 和 TR1 显示出对氧化应激的反应上调 [36],当硒供应受到限制时,这种反应更为明显。值得一提的是,上述反应还取决于氧化应激的水平,因为在轻度氧化应激中确实如此,但在极高水平时,会激活其他一些机制[2]。
很可能在细胞培养中,这些硒蛋白的表达/活性增加是对纳米硒产生的氧化应激的反应,而不是硒供应改善的反映。因此,在解释基于细胞培养研究的结果时应该谨慎。最近的研究表明,生物源性纳米硒可以激活核因子(红系衍生 2)样 2(Nrf2)并增加其下游基因的表达,负责以剂量和时间依赖性方式合成抗氧化剂 [37]。此外,作者发现Nrf2的敲低显着阻断了这种纳米Se颗粒的抗氧化作用。
在批判性地分析补充纳米硒对家禽、农场动物和鱼类的生产和繁殖性能的积极影响时,有必要考虑到,在大多数情况下,硒并没有提高性能,而是防止了由于环境或营养限制。在许多情况下,动物实验是在极低的硒背景水平下进行的,因此,以任何可用的形式添加硒都会产生积极的影响。
将纳米硒商业化用作饲料添加剂的挑战可能如下:
- 1.
有必要了解纳米硒在细胞、亚细胞和基因水平上的吸收、同化和作用的分子机制。如果没有这些数据,就很难获得可重复的结果并找到对观察到的影响的正确解释。例如,在最近发表的一篇论文中,由于鸡体内纳米硒过量 [38],肝脏中只有少数基因(18 个蛋白质,没有一个与硒代谢直接相关)受到影响,而其他膳食硒的形式,包括SeMet,可以影响数百个基因。
- 2.
了解纳米硒是否可以像 SeMet 一样在体内建立任何硒储备,以及这些储备是否在压力条件下可用,这一点很重要。
- 3.
有必要设计一种能够提供相同尺寸、稳定性和相当好的(至少 6-12 个月)可储存性的纳米硒颗粒的技术[2]。
- 4.
微生物益生菌很可能可用于转化用作饲料添加剂的纳米硒。一方面,这种益生菌可以包含有助于肠道中纳米硒同化的特定微生物(例如韦隆氏菌 物种)。另一方面,富含硒的益生菌可能是另一种可能性[34, 39]
- 5.
应全面解决副作用、风险和环境问题。事实上,需要进一步研究以告知政策制定者和监管机构有关纳米硒的纳米毒理学潜力 [40]。特别是,干燥形式的非常小的颗粒使产品非常多尘,并且异常的纳米硒颗粒行为一旦进入体内,就会发出警告,主要关注饲料厂工人的保护。由于肠道微生物群通过甲基化和元素硒的形成负责排出过量的硒[34],微生物群的增强/调节可以为应对可能的纳米硒毒性开辟新的视野。
- 6.
纳米硒在动物营养中的积极作用不应掩盖其使用可能带来的不利后果。事实上,纳米粒子在各种条件下的行为可能会有很大差异,在我们了解如何控制这种行为之前,纳米硒在广泛的工业规模上的使用应该是不可能的。例如,在考虑纳米硒吸收时,有必要提及所谓的特洛伊木马效应,即纳米颗粒可能对肠道中的其他物质具有渗透增强特性 [41]。这可能会产生一些问题,因为饲料中有一系列“不需要的”化合物,而肠道可以防止它们被吸收。事实上,纳米硒在肠道中的行为值得进一步研究。
- 7.
在上述综述 [1] 中,纳米硒的抗菌和抗癌特性得到了极大的关注,纳米硒的独特特性似乎有助于对抗各种疾病。事实上,纳米硒可以被认为是一种用于各种医疗条件的新药,包括癌症治疗,而其作为有效饲料添加剂的用途则颇受质疑。未来的研究必须回答这些重要的问题和担忧。
结论
在过去的几年里,发表了一系列关于纳米硒的论文,并且该主题正在迅速发展。然而,在这种形式的硒找到商业家禽/动物生产的方法之前,有必要了解和解释纳米硒如何转化为活性硒蛋白。纳米硒作用的一种可能机制/途径可能是由肠道微生物群介导的,肠道微生物群可以将纳米硒转化为亚硒酸盐、H2Se 或硒磷酸盐,并合成以下硒蛋白(图 1)。有证据表明,在自然界中,细菌可以还原或氧化金属硒(Se 0 ) 产生 Se −2 或 Se +4 和Se +6 分别。在参与硒氧化还原变化的微生物中,Veillonella 属 值得特别关注,因为食用动物(包括鸡肉)的肠道中存在此类细菌。肠道微生物群参与纳米硒同化和代谢的可能性有待进一步研究,纳米硒作为硒源在动物/家禽营养中的优缺点有待进一步的批判性评估。
纳米材料
- DevOps 和云相互需要的 3 个原因; DevCloudOps
- PVDF 螺钉、六角螺栓、六角螺母、其他紧固件和定制零件
- Ultem® PEI 螺钉、六角螺栓、定制零件和其他紧固件
- 通过生物育种获得的植物和动物在 EPO 不再具有专利权
- 3 机器人仿真对制造商和其他机器人用户的优势
- SAP S/4HANA® 中的流程自动化 - 希望、驱动因素和行动
- Nexus Integra 与其他物联网和大数据平台的区别
- PCB工程师使用什么软件和其他工具?
- 镍 718 成分和其他关键合金
- 使用铝金属残余物、水滴和其他金属短裤的优点
- 最重要的货运安全问题以及如何避免它们
- 卧式加工中心 (HMC) – 价格、成本和其他考虑因素