

人类脐带间充质干细胞衍生的外泌体 MicroRNA-18b-3p 通过靶向 LEP 抑制先兆子痫的发生
摘要
源自表达 microRNA 的人脐带间充质干细胞 (hucMSCs) 的外泌体已在人类疾病中得到强调。然而,hucMSCs 衍生的外泌体 miR-18b-3p 对先兆子痫(PE)的详细分子机制仍有待进一步研究。我们旨在研究外泌体和 miR-18b-3p/leptin (LEP) 对 PE 发生的影响。鉴定了hucMSC和hucMSC-外泌体(Exos)的形态。外泌体感染不同的表达 miR-18b-3p 的慢病毒,以探索 miR-18b-3p 在 PE 中的作用。腹腔注射N建立PE大鼠模型 -硝基-L-精氨酸甲酯。在 PE 大鼠胎盘组织中测试了 LEP 和 miR-18b-3p 的表达。此外,检测到外泌体对 LEP 和 miR-18b-3p 表达的影响。检测PE大鼠的收缩压(SBP)、蛋白尿、炎症因子、胎鼠和胎盘重量以及细胞凋亡情况。最后,使用双荧光素酶报告基因测定和 RNA 下拉测定验证了 miR-18b-3p 和 LEP 之间的关系。外泌体、恢复miR-18b-3p或抑制LEP可降低PE大鼠的收缩压和蛋白尿,增加胎鼠和胎盘的重量,降低血清炎症因子水平,抑制PE大鼠的凋亡细胞,发挥抑制作用关于 PE 进展。 PE大鼠胎盘组织中miR-18b-3p降低,LEP升高。 LEP是miR-18b-3p的直接靶基因。 miR-18b-3p 的上调或外泌体的处理抑制了 LEP 表达并减少了 PE 的发生,而 miR-18b-3p 的下调具有相反的效果。下调的 LEP 逆转了 miR-18b-3p 减少对 PE 大鼠的影响。 HucMSCs 来源的外泌体 miR-18b-3p 靶向 LEP 参与 PE 的发生和发展。该研究可为PE的机制及研究提供新的理论依据。
介绍
先兆子痫 (PE) 以蛋白尿和高血压为特征 [1],是人类妊娠期胎儿和孕产妇死亡和发病的主要原因 [2]。 PE 的病因和发病机制尚不清楚 [3],据报道,PE 与异常滋养细胞侵袭导致母体内皮功能障碍、慢性胎盘灌注不良和高血压有关,从而导致不良后果 [4]。除了胎儿和胎盘分娩外,没有针对 PE 的特定疗法 [5]。因此,迫切需要探索治疗靶点以改善该病的预后。
人脐带 (huc) 是间充质干细胞 (MSC) 的合适来源,其分泌多种营养因子和细胞因子,并显示出强大的抗炎和免疫调节能力 [6]。一项研究证实,PE 可加速脐带沃顿氏果冻衍生的 MSCs 中神经胶质标志物的表达 [7]。也有报道称,hucMSCs 外泌体 (Exos) 对 PE 大鼠的胎盘形态和血管生成具有保护作用 [8]。外泌体是小的 (50-100 nm) 分泌性囊泡,通过包裹致癌因子并将致癌因子通过循环传递到远处或周围细胞,从而介导肿瘤微环境中细胞之间的通讯 [9]。一项研究揭示了通过外泌体将 sFlt-1 和 sEng 有效转移到 PE 患者的内皮细胞中引起的血管功能损伤和并发症 [10]。 MicroRNA (miRNA) 是内源性非编码 RNA,长度为 18-25 个核苷酸,在转录后水平调节基因的表达 [11]。一项研究中的数据报道,miR-18b 表达影响 PE 中滋养层细胞的细胞侵袭、活力和迁移 [12]。此外,吴等人。已经提出 miR-18b 减弱高糖诱导的人视网膜内皮细胞的增殖,这可能为理解糖尿病视网膜病变的发病机制提供新的见解 [13]。然而,hucMSC 衍生的外泌体 miR-18b-3p 在 PE 中的作用仍然未知。瘦素 (LEP) 对细胞分化/增殖和生理状态的免疫具有多效作用,除胎盘等其他组织外,主要来自脂肪细胞 [14]。一项研究证实,异常的 LEP 启动子甲基化与 PE 进展有关 [15]。另一项研究表明,胎盘是妊娠期 LEP 表达的主要部位 [16]。尽管如此,miR-18b-3p 和 LEP 之间的结合关系仍然难以捉摸。因此,我们旨在探讨hucMSC衍生的外泌体miR-18b-3p在LEP参与的PE中的作用,我们推断hucMSC衍生的外泌体miR-18b-3p可能通过靶向LEP抑制PE进展。
材料和方法
道德批准
该研究得到了武汉大学人民医院机构审查委员会的批准。所有参与者都签署了知情同意书。所有动物实验均符合武汉大学人民医院国际委员会《实验动物饲养和使用指南》。
HucMSCs 的分离、培养和鉴定
收集健康产妇分娩的胎儿脐带,切碎,过筛,与磷酸盐缓冲液(PBS)混合。脐带组织以 1500 r/min 离心 5 分钟,离心半径为 10 厘米。将组织用含有 10% 胎牛血清 (FBS) 的 Dulbecco 改良 Eagle 培养基 (DMEM)/F12 悬浮并转移到培养瓶中。 4 天后换液,然后每 3 天换液一次。当汇合度达到90%左右时进行细胞传代培养。在光学显微镜下观察hucMSCs的贴壁生长和形态。细胞用油红O染色液(Beyotime Institute of Biotechnology, Shanghai,China)染色检测hucMSCs的成骨分化,用碱性磷酸酶(ALP)染色液(Beyotime)染色检测hucMSCs的成脂分化。采用流式细胞仪(Beckman Coulter Life Sciences,Brea,CA,USA)测试 CD73、CD166(均为 1:10,BD Biosciences,Franklin Lakes,NJ,USA)和 CD105(1:20,AbD Serotec,Oxford,英国)。
HucMSC-Exos的提取与鉴定
培养生长良好的hucMSCs。收集上清液并以 10 cm 离心半径以 28,500 r/min 离心 1 小时。弃上清,细胞用2%戊二醛和1%锇酸固定,乙醇脱水,环氧丙烷浸渍,干燥2小时,Epon812包埋,切片。切片分别用铀和铅染色。最后,在电子显微镜下观察外泌体。 Nanosight 检测器(Malvern Instruments,Malvern,UK)用于检测外泌体纳米粒子的布朗运动成像及其大小。通过蛋白质印迹法鉴定hucMSC-Exos的表面标志物,结果显示hucMSC-Exos表达CD9、CD81和CD63。
慢病毒感染方法
用含有低表达 miR-18b-3p 载体和低表达 miR-18b-3p 载体阴性对照(NC)的慢病毒感染 HucMSC(上海基因制药有限公司,上海,中国)。最终得到稳定表达的hucMSC-antagomir NC和hucMSC-miR-18b-3p antagomir。细胞培养48 h,收集上清液,超速离心得到相应的Exos-antagomir NC和Exos-miR-18b-3p antagomir。
实验动物
选择健康清洁水平和性成熟的Wistar大鼠(体重200-250 g,年龄8 w,不分性别)(武汉大学实验动物中心,武汉,中国)。大鼠在屏障系统中饲养,温度为 18-28°C,相对湿度为 40-70%,食物和水充足。
大鼠PE模型的建立
参考文献[17]腹腔注射50mg/kg一氧化氮合成酶抑制剂N(G)-硝基-l-精氨酸甲酯(L-NAME, Beyotime)建立大鼠PE模型。 PE模型的成功建立基于血压升高20 mmHg和高于115 mmHg以及蛋白尿增多。
动物分组
雌性大鼠和雄性大鼠按 1:1 随机同居,下午 5-6 点将两只大鼠单独饲养在一个特殊的笼子里。前一天。次日用阴道塞和显微镜观察雌性大鼠阴道分泌物中的精子。如果同时结果为阳性,则记录为妊娠第 0 天。从 13 rd 妊娠当天,大鼠分为6组(每组10只):正常组(从妊娠第13天至第20天腹腔注射等量生理盐水),PE组(L-NAME [50 mg/ kg/天]在妊娠第13天至第20天腹腔注射,在妊娠第16天至第19天向胎盘注射20μL生理盐水),PE + miR-NC组(L-NAME [50 mg /kg/天]在妊娠第 13 天至第 20 天腹腔注射,在妊娠第 16 天至第 19 天向胎盘注射 20 μL 4 nmol miR-NC),PE + miR-18b-3p agomir组(L-NAME [50 mg/kg/天] 在妊娠第 13 天至第 20 天腹腔注射,并在妊娠第 16 天至第 19 天向胎盘注射 20 μL 4 nmol miR-18b-3p agomir) , PE + miR-18b-3p antagomir组(L-NAME [50 mg/kg/天]从妊娠第 13 天到第 20 天腹腔注射,20 μL 4 nmol miR-18b-3p antagomir 注射到p妊娠第 16 天至第 19 天的胎盘),PE + miR-18b-3p antagomir + 小干扰 RNA (si)-LEP 组(L-NAME [50 mg/kg/天] 从第 13 天至第 20 天腹膜内注射)在妊娠第 16 天至第 19 天将 20 μL 4 nmol miR-18b-3p antagomir 和 si-LEP 注射到胎盘中)和 PE + si-LEP 组(L-NAME [50 mg/kg per天]从妊娠第 13 天至第 20 天腹膜内注射,并在妊娠第 16 天至第 19 天向胎盘注射 20 μL 4 nmol si-LEP)。用外泌体和携带慢病毒的外泌体治疗大鼠。将大鼠分为 5 组(每组 10 只大鼠):正常组(从妊娠第 13 天至第 20 天腹腔注射等量生理盐水)、PE 组(L-NAME(每天 50 mg/kg) ),在妊娠第 13 天至第 20 天腹腔注射,在妊娠第 16 天至第 19 天向胎盘注射 20 μL 生理盐水),PE + Exos 组(L-NAME(每天 50 mg/kg)从妊娠第 13 天到第 20 天腹腔注射,在妊娠第 16 天到第 19 天向胎盘注射 20 μL Exos(80 μg 外泌体悬浮在 20 μL 生理盐水中),PE + Exos-antagomir NC组(L-NAME(50 mg/kg/天)在妊娠第 13 天至第 20 天腹腔注射,20 μL Exos-antagomir NC(80 μg 外泌体悬浮在 20 μL 生理盐水中)注射到胎盘在妊娠第 16 天至第 19 天)和 PE + Exos-miR-18b-3p antagomir 组(L-NAME(每天 50 mg/kg)从第 13 天开始腹膜内注射)至妊娠第 20 天,在妊娠第 16 天至第 19 天向胎盘注射 20 μL Exos-miR-18b-3p antagomir(80 μg 外泌体悬浮于 20 μL 生理盐水中)。
收缩压 (SBP) 的检测和 24 小时蛋白尿的测定
大鼠血压采用大鼠尾动脉血压测量法。在妊娠第 10、13、16 和 19 天使用大鼠尾动脉压力检测器(Tensys (R) Medical Inc., San Diego, CA, USA)测量所有妊娠大鼠的尾套收缩压。短时间内测3次压力;然后取平均值作为血压值。
在自由饮食和饮水的情况下,于妊娠第10、13、16、19天采集孕鼠24小时尿,在武汉大学人民医院肾内科检测蛋白质含量。
示例集合
妊娠大鼠在妊娠第 21 天用 3% 戊巴比妥钠麻醉。大鼠外周血保存,离心取血清,-20℃冰箱保存备用。然后剖宫产取胎鼠和胎盘,去除胎膜和相连的脐带,切断与胎鼠相连的脐带。将胎盘和胎鼠置于无菌纱布上分别擦干血液和羊水,然后放在分析天平上称重。一部分胎盘组织用4%多聚甲醛固定,乙醇脱水,二甲苯清除,石蜡包埋,连续交叉切片(5μm)进行苏木精-伊红(HE)染色和末端脱氧核苷酸转移酶介导的脱氧尿苷三磷酸-生物素缺口末端标记 (TUNEL) 染色。其余保存于-80℃,用于逆转录定量聚合酶链反应(RT-qPCR)检测、蛋白质印迹分析和酶联免疫吸附试验(ELISA)。
ELISA
ELISA检测血清中肿瘤坏死因子-α(TNF-α)、白细胞介素(IL)-1β和IL-6的含量。按照试剂盒(R&D Systems, Minneapolis, MN, USA)的说明测定TNF-α、IL-1β和IL-6的浓度。通过酶标仪(Thermo Fisher Scientific,MA,USA)测试光密度(OD)值(490 nm)。以OD值为横坐标,以相应标准样品的浓度为纵坐标,得到相应的标准曲线。根据标准曲线计算TNF-α、IL-1β和IL-6浓度。
HE 染色
胎盘组织石蜡样品二甲苯澄清,常规梯度酒精脱水,苏木精染色,1%盐酸酒精鉴别,1%氨水返蓝。组织用1%伊红溶液复染,脱水(分别为75%、90%、95%乙醇,无水乙醇),二甲苯透明,干燥,封闭,电镜观察。
TUNEL 染色
石蜡包埋切片按说明书常规脱蜡脱水,然后用TUNEL试剂盒(南京科金生物科技有限公司,江苏,中国)检测细胞凋亡。 4,6-二氨基-2-苯基吲哚(上海百泰生物科技有限公司,中国上海)用荧光显微镜(Nikon,Tokyo,Japan)观察TUNEL阳性细胞[18]。
RT-qPCR
称重胎盘组织。每 50-100 mg 胎盘组织中加入 1 mL TRIzol(Invitrogen,Carlsbad,California,USA)并完全溶解。在组织中加入 200 μL 氯仿,并在 4°C、12,000 rpm 下离心以提取总 RNA。 RNA的浓度和纯度由DU-800蛋白质核酸分光光度计(Beckman)测定。 U6 和 β-肌动蛋白用作加载对照。 PCR引物由上海生工生物技术有限公司(中国上海)设计和合成。引物序列见表1。根据RNA逆转录试剂盒(Sangon)的说明将RNA逆转录为cDNA。 PCR扩增产物经琼脂糖凝胶电泳验证。数据由 2 −ΔΔCt 计算 方法。
蛋白质印迹分析
胎盘组织总蛋白通过放射免疫沉淀法细胞裂解缓冲液(Beyotime)提取。 HucMSC-Exo 用于提取缓冲液,以 14,000 rpm 离心。保留上清液用于检测血清中外泌体标志蛋白(CD81、CD63和CD9)的蛋白表达。通过二辛可宁酸试剂盒(Beyotime,P0010)测定蛋白质浓度。根据蛋白质定量结果上样,用10%十二烷基硫酸钠-聚丙烯酰胺凝胶电泳处理,转膜。膜用 5% 脱脂牛奶封闭,用初级抗体 LEP、CD63、CD81、CD9 和 β-肌动蛋白(4 mL,1:1000,Santa Cruz Biotechnology, Inc, Santa Cruz, CA, USA)探测,重新探测用 4 mL 二抗山羊抗兔 IgG/辣根过氧化物酶,暴露和显影。 β-肌动蛋白用作内参。灰度值通过凝胶图形分析软件Image Lab进行分析。
双荧光素酶报告基因检测
采用在线预测软件https://cm.jefferson.edu/预测miR-18b-3p与LEP的靶标关系以及miR-18b-3p与LEP 3′非翻译区(UTR)的结合位点)。组成含有 miR-18b-3p 结合位点的 LEP 3'UTR 启动子区序列。构建了 LEP 3'UTR 野生型 (WT) 质粒和 LEP 3'UTR 突变型 (MUT)。重组质粒分别命名为LEP 3'UTR-WT和LEP 3'UTR-MUT。将培养的 293T 细胞与 miR-18b-3p 模拟物和 LEP 3'UTR-WT、miR-18b-3p 模拟物和 LEP 3'UTR-MUT、模拟物 NC 和 LEP 3'UTR-WT、模拟物 NC 和LEP 3'UTR-MUT 30 小时。然后收集293T细胞。根据双荧光素酶报告基因检测试剂盒(Promega,Madison,WI,USA),通过发光测量测定细胞中萤火虫和海肾荧光素酶活性。
RNA 下拉分析
生物素化的 RNA 探针(Bio-miR-NC、Bio-miR-18b-3p 和 Bio-miR-18b-3p-Mut)与 293T 细胞的裂解物一起孵育,并使用与抗生素链霉素缀合的磁珠提取。实验根据 Pierce 磁性 RNA 下拉试剂盒(Pierce,IL,USA)的说明进行。使用 TRIzol (Pierce) 洗脱和纯化 RNA。如前所述[19],使用RT-qPCR定量RNA复合物中LEP的富集。
统计分析
所有数据均由 SPSS 21.0 软件(IBM Corp. Armonk, NY, USA)解释。测量数据表示为平均值 ± 标准偏差。数据由独立样本t进行 检验两组比较,而多组之间的比较通过单向方差分析 (ANOVA) 和 Tukey 事后检验进行评估。统计显着性的标准设置为 p <0.05.
结果
HucMSC 和 HucMSC-Exos 的形态和鉴定
倒置显微镜下观察脐带组织肿块。可以看到第3天细胞从组织块中爬出;细胞显示纺锤形和细丝,并在大约 5 天时像集落一样生长。培养至第3代细胞形态为均一的长梭形,与成纤维细胞形态相似,排列规则(图1a)。 HucMSC成脂分化2w后,细胞质内形成脂滴,倒置显微镜下脂滴呈Kranz结构(图1b),表明分离培养的hucMSC具有成脂分化能力。成骨分化2 w后,倒置显微镜下可见大量棕色钙质结节(图1c),表明分离培养的hucMSC具有成骨分化能力。采用流式细胞仪检测细胞免疫表型,结果显示细胞高表达MSCs表面标志物CD73、CD105和CD166(图1d)。
<图片>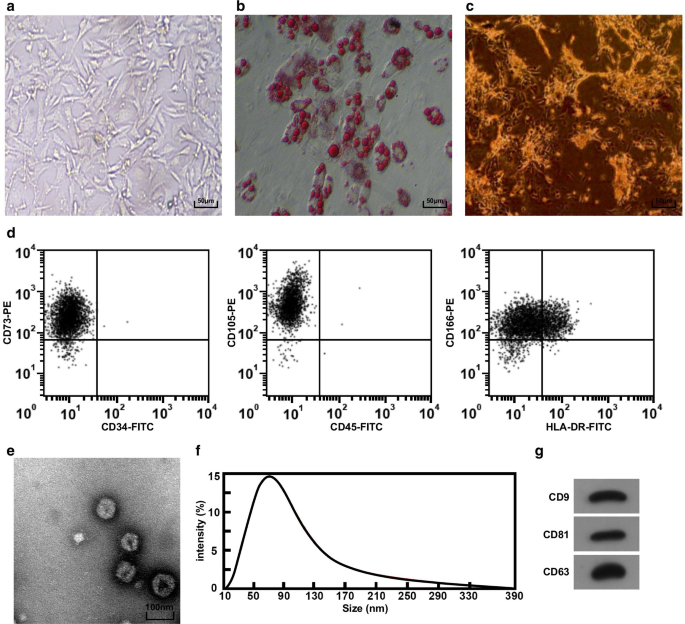
HucMSC 和 HucMSC-Exos 的形态学和鉴定。 一 倒置显微镜下hucMSC的形态,b 通过油红O染色测试hucMSC。 c 通过ALP染色测试hucMSC。 d 流式细胞术用于检测免疫表型。 e 通过 TEM 观察到的 hucMSC-Exos 的形状和大小。 f 使用 Nanosight 分析检测外泌体的粒径分布。 g Western blot法检测hucMSC-Exos中CD9、CD81和CD63蛋白表达
通过TEM观察hucMSC-Exos的形态,结果显示外泌体呈圆形或椭圆形,中心密度低,两侧染色较厚(图1e)。 Nanosight分析用于分析外泌体的粒径,结果表明粒径主要分布在40-100 nm之间,80 nm附近较为集中(图1f)。 Western印迹分析显示所有表面标志物CD81、CD63和CD9均在hucMSC-Exos中表达(图1g)。
恢复 miR-18b-3p 减轻 PE 大鼠的病理特征
SBP和24h蛋白尿结果显示:给药前(妊娠第10天)6组SBP和24h蛋白尿无显着差异。妊娠第19天的收缩压和24小时蛋白尿与正常大鼠无明显差异。在 PE 大鼠或用 miR-NC、miR-18b-3p agomir、miR-18b-3p antagomir、miR-18b-3p antagomir + si-LEP、si-NC 或 si-LEP、SBP 和 24-h 治疗的 PE 大鼠或 PE 大鼠中蛋白尿在妊娠第 13 天开始增加。用miR-18b-3p agomir和si-LEP治疗的PE大鼠在妊娠第16天和第19天的SBP和24小时蛋白尿没有明显差异。 PE 大鼠在妊娠第 19 天 SBP 和 24 小时蛋白尿增加;这种增加因 miR-18b-3p 升高而减少,但通过 miR-18b-3p 抑制进一步增强; LEP 降低消除了 miR-18b-3p 下调在 PE 大鼠妊娠第 19 天 SBP 和 24 小时蛋白尿中的作用(图 2a、b)。
<图片>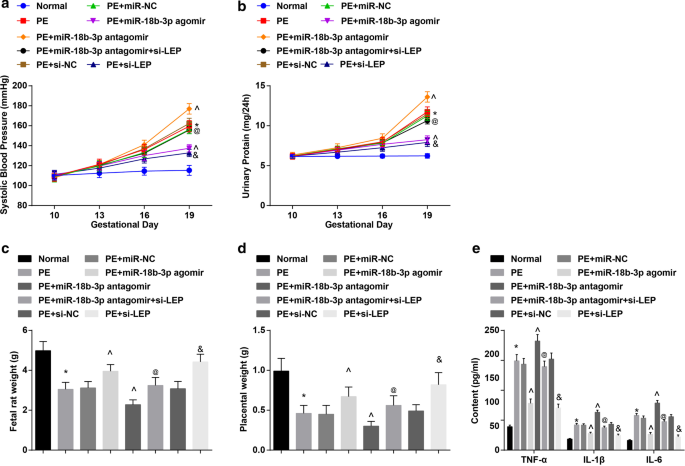
恢复miR-18b-3p可减轻PE大鼠的病理特征。 一 大鼠 SBP 的结果。 b 大鼠 24 小时蛋白尿的结果。 c 胎鼠体重变化。 d 大鼠胎盘重量的变化。 e ELISA检测血清炎症因子的变化。 n =10, *p <0.05 与正常组相比。 ^p <0.05 与 PE + miR-NC 组相比。 @ p <0.05 与 PE + miR-18b-3p antagomir 组相比。 & p <0.05 与 PE + si-NC 组相比。测量数据以均值 ± 标准差表示,多组间比较采用单向方差分析和Tukey检验
PE大鼠胎鼠和胎盘重量减轻;上调 miR-18b-3p 或下调 LEP 增加,而下调 miR-18b-3p 降低 PE 大鼠胎鼠和胎盘的重量; LEP沉默逆转了miR-18b-3p抑制对PE大鼠胎鼠和胎盘重量的影响(图2c,d)。
检测PE大鼠血清中的炎症因子。发现PE大鼠体内TNF-α、IL-1β和IL-6含量升高; miR-18b-3p升高或LEP抑制受到抑制,而miR-18b-3p降低促进TNF-α、IL-1β和IL-6的含量; LEP耗竭消除了抑制的miR-18b-3p对TNF-α、IL-1β和IL-6含量的影响(图2e)。
过表达的 miR-18b-3p 改善 PE 大鼠胎盘组织的组织病理学变化
正常大鼠胎盘绒毛血管丰富,结构清晰,胎盘绒毛中以合体滋养细胞为主,细胞滋养细胞较少。 PE大鼠或用miR-NC、miR-18b-3p antagomir、si-NC或miR-18b-3p antagomir + si-LEP治疗的PE大鼠,胎盘绒毛数量减少,结构模糊萎缩,部分绒毛进行纤维蛋白样坏死,胎盘绒毛合体滋养细胞结节数增多,大部分绒毛未成熟。 miR-18b-3p agomir和si-LEP治疗的PE大鼠滋养细胞数量减少,病理变化减轻(图3a)。
<图片>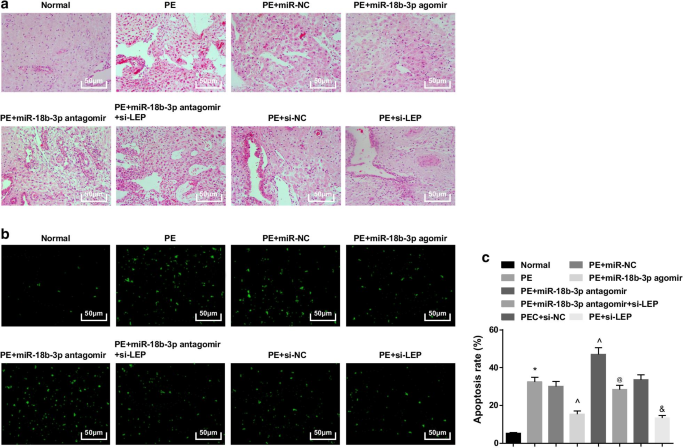
过表达的 miR-18b-3p 可改善 PE 大鼠胎盘组织的病理变化并抑制其凋亡。 一 HE染色用于检测胎盘组织的病理特征。 b 实施TUNEL染色以测定PE大鼠胎盘组织的凋亡细胞。 c TUNEL染色检测细胞凋亡率。 n =10, *p <0.05 与正常组相比。 ^p <0.05 与 miR-NC 组相比。 @ p <0.05 与 miR-18b-3p antagomir 组相比。 & p <0.05 与 si-NC 组相比。测量数据以均值 ± 标准差表示,多组间比较采用单向方差分析和Tukey检验
TUNEL染色提示可见少量凋亡细胞。 PE大鼠的凋亡细胞增加,miR-18b-3p升高和LEP沉默减少,miR-18b-3p抑制进一步增强; LEP沉默还逆转了miR-18b-3p抑制对PE大鼠凋亡细胞数量的影响(图3b、c)。
总之,miR-18b-3p上调或LEP抑制的大鼠在组织学上PE进展程度降低,而沉默的LEP可以消除抑制miR-18b-3p的治疗效果。
miR-18b-3p 下调,而 LEP 在 PE 大鼠胎盘组织中上调,miR-18b-3p 靶向 LEP
基于以上结果,LEP下调逆转了miR-18b-3p下调对PE大鼠病理学和组织学的治疗效果;因此,我们假设miR-18b-3p可能与LEP有关。
蛋白质印迹分析和 RT-qPCR 显示 PE 大鼠降低了 miR-18b-3p 并增加了 LEP 表达水平; miR-18b-3p agomir的治疗在PE大鼠中上调miR-18b-3p并下调LEP,而miR-18b-3p antagomir的治疗增加了LEP表达; LEP沉默逆转了miR-18b-3p减少对PE大鼠LEP表达的促进作用(图4a-c)。
<图片>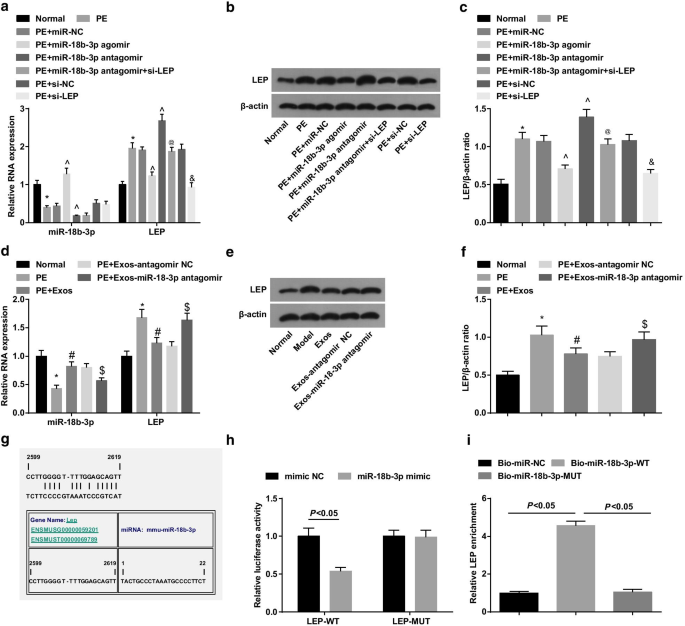
PE大鼠胎盘组织中miR-18b-3p下调,LEP上调。 一 使用RT-qPCR检测胎盘组织中miR-18b-3p和LEP mRNA的表达。 b 胎盘组织中 LEP 的蛋白条带。 c 进行蛋白质印迹分析以检测胎盘组织中的LEP蛋白表达。 d 使用RT-qPCR检测外泌体处理后胎盘组织中miR-18b-3p和LEP mRNA的表达。 e 外泌体处理后胎盘组织中LEP的蛋白条带。 f 进行蛋白质印迹分析以检测外泌体处理后的 LEP 蛋白表达。 g 由在线软件预测的 miR-18b-3p 和 LEP 的结合位点。 h 双荧光素酶报告基因检测证实miR-18b-3p与LEP之间的靶标关系。 我 miR-18b-3p 和 LEP 之间的靶向关系通过 RNA pull-down 测定验证。 n =10, *p <0.05 与正常组相比。 ^p <0.05 与 miR-NC 组相比。 @ p <0.05 与 miR-18b-3p antagomir 组相比。 & p <0.05 与 si-NC 组相比。 # p <0.05 与 PE 组相比。 $ p <0.05 与 PE + Exos-antagomir NC 组相比。测量数据以均值 ± 标准差表示,多组间比较采用单向方差分析和Tukey检验
使用蛋白质印迹分析和 RT-qPCR 来探索外泌体在 PE 大鼠中的作用。结果显示外泌体在 PE 大鼠中上调 miR-18b-3p 并下调 LEP,表明外泌体对 PE 发展有抑制作用。此外,携带miR-18b-3p antagomir的外泌体在PE大鼠中诱导miR-18b-3p下调和LEP上调(图4d-f)。
miR-18b-3p 和 LEP 之间的靶标关系由生物信息学在线预测软件 https://cm.jefferson.edu/ 预测(图 4g)。双荧光素酶报告基因分析表明,miR-18b-3p 模拟物降低了 LEP 3'UTR-WT 的荧光素酶活性,而对 LEP 3'UTR-MUT 的荧光素酶活性没有影响(图 4h)。此外,RNA pull-down 测定表明,WT 生物素化的 miR-18b-3p 增加了 LEP 富集(图 4i)。这些发现表明LEP是miR-18b-3p的靶基因。
hucMSC-Exos 减弱 PE 大鼠的病理特征
The results of SBP and 24 h presented that there was no significant difference in SBP and 24-h proteinuria in 5 groups before administration (day 10 of gestation). SBP and 24-h proteinuria in day 19 of gestation showed no distinct difference in normal rats. In PE rats, SBP and 24-h proteinuria began to raise at day 13 of gestation. There was no distinct difference of SBP and 24-h proteinuria in day 16 and day 19 of gestation in PE rats treated with hucMSC-Exos and hucMSC-Exos transmitting antagomir NC. SBP and 24-h proteinuria heightened in day 19 of gestation in the PE rats, while the increase was reduced by injection of hucMSC-Exos. Inhibiting miR-18b-3p reversed the effect of hucMSC-Exos on SBP and 24-h proteinuria in day 19 of gestation in PE rats (Fig. 5a, b).

hucMSC-Exos attenuate pathological characteristics of PE rats. 一 Results of SBP in rats after exosome treatment. b Results of 24-h proteinuria in rats after exosome treatment. c Weight changes of fetal rats after exosome treatment. d Changes of placental weight in rats after exosome treatment. e Changes of inflammation factors after exosome treatment in serum were determined using ELISA. n = 10, *p < 0.05 versus the normal group. # p < 0.05 versus the PE group. $ p < 0.05 versus the PE + Exos-antagomir NC group. Measurement data were depicted as mean ± standard deviation, and comparisons among multiple groups were assessed by one-way ANOVA followed by Tukey’s test
The weight of fetal rat and placenta was measured, and we found that the PE rats had decreased weight of fetal rat and placenta; miR-18b-3p downregulation abolished the role of hucMSC-Exos in the weight of fetal rat and placenta in PE rats (Fig. 5c, d).
Inflammatory factors in serum were detected using ELISA. TNF-α, IL-1β and IL-6 contents remarkably increased in PE rats. Exosomes treatment decreased TNF-α, IL-1β and IL-6 contents in serum of PE rats, which were enhanced by injection of exosomes inhibiting miR-18b-3p (Fig. 5e).
Exosomes Alleviates Pathological Change and Inhibit Apoptosis of Placenta Tissues of PE Rats
In normal rats, the placental villus was abundant in blood vessels with a clear structure, syncytiotrophoblasts were the main trophoblast in placental villi, and there were fewer cytotrophoblasts. In the PE rats and PE rats treated with hucMSC-Exos-miR-18b-3p-antagomir, the number of placental villi reduced, the structure was blurred and atrophied, some villi were presented fibrinoid necrosis, and the number of syncytiotrophoblast nodules in placental villi enhanced, and most of the villi were immature. The pathological change was improved in the PE rats treated with hucMSC-Exos or hucMSC-Exos-antagomir NC versus the PE rats and PE rats treated with hucMSC-Exos-miR-18b-3p antagomir (Fig. 6a).

Exosomes alleviate pathological change and decrease apoptotic cells of placenta tissues in PE rats. 一 HE staining was utilized to test pathological features of placenta tissues in PE rats after exosome treatment. b TUNEL staining was implemented to determine apoptotic cells of placenta tissues in PE rats after exosome treatment. c Cell apoptosis rate was detected by TUNEL staining. n = 10, *p < 0.05 versus the normal group. # p < 0.05 versus the PE group. $ p < 0.05 versus the PE + Exos-antagomir NC group. Measurement data were depicted as mean ± standard deviation, and comparisons among multiple groups were assessed by one-way ANOVA followed by Tukey’s test
TUNEL staining indicated that in normal rats, a small number of apoptotic cells could be seen. PE rats had enhanced apoptotic cells, and reduced miR-18b-3p reversed the impacts of hucMSC-Exos on the number of apoptotic cells in placenta tissues from PE rats (Fig. 6b, c).
Discussion
PE is a multisystem pregnancy disorder characterized by proteinuria and either high blood pressure or other adverse conditions and is linked to a wide range of maternal endothelial dysfunction [20]. It was reported that hucMSC-Exo improved the morphology of placental tissue in PE rats through suppressing cell apoptosis and facilitating angiogenesis in placental tissue in a dose-dependent manner [8]. A study has reported that miR-18b expression affected cell migration, viability and invasion in PE [12]. Moreover, it was verified increased maternal LEP concentration and hypomethylation of the LEP in placenta in early onset PE [21]. The current study was designed to explore the effect of exosomes and miR-18b-3p targeted LEP on the occurrence of PE. The findings in this study revealed that hucMSC-derived exosomal miR-18b-3p inhibited PE progression by reducing LEP.
Based on our findings, miR-18b-3p reduced and LEP elevated in placenta tissues of PE rats. Similar to our study, the mRNA expression of miR-18b was markedly suppressed in PE placental tissues relative to that in normal placental tissues [12]. In addition, a study revealed that miR-18b content was dramatically reduced in malignant melanoma tissues in comparison with their matched adjacent non-tumor tissues [22]. Another study has verified that placental LEP expression was raised in preterm PE compared with controls [23]. Moreover, a study showed that LEP expression was obviously heightened in preeclamptic placentas [15]. This literature provided a theoretical basis for us to explore the abnormal expression of miR-18b-3p and LEP in PE. Moreover, it was predicted using a bioinformatic software that LEP was targeted by miR-18b-3p, and this targeting relationship was further confirmed with dual-luciferase reporter gene assay in our research. A study reported that LEP is a target for all three miRNAs (miR-1301, miR-223 and miR-224) in early-onset PE [16]. Another study has displayed that LEP decreased miR-93 expression in osteoarthritis and rheumatoid arthritis [24]. However, the binding between miR-18b-3p and LEP in human diseases, especially in PE, remains scarcely studied, which is the novelty of this study. Furthermore, a result emerging from our study reported that exosomes increased miR-18b-3p and decreased LEP in placenta tissues of PE. It was formerly documented that the expression of miR-18b-5p was notably raised in colorectal cancer plasma exosomes [25], while the relationship between hucMSC-Exos and miR-18b-3p/LEP in PE needs further study.
Additionally, the finding from our investigation showed that restored miR-18b-3p reduced SBP and 24-h proteinuria of PE rats, increased the weight of placenta, declined TNF-α, IL-1β and IL-6 contents in serum and placenta tissues as well as suppressed cell apoptosis. These data indicated that miR-18b-3p elevation contributes to alleviating the symptoms and pathological changes in PE. It was demonstrated that stable upregulation of miR-18b produced effective tumor inhibitor activity, such as inhibiting melanoma cell viability, inducing apoptosis and reducing tumor growth in vivo [26]. Another result in our study was that hucMSC-Exos reduced SBP and 24-h proteinuria of PE rats, increased the weight of placenta, declined TNF-α, IL-1β and IL-6 contents in serum and placenta tissues as well as suppressed cell apoptosis. The findings of the current study revealed that exosomes treated PE rat models presented an increase of the number and quality of fetuses, the quality of placenta, but cell apoptosis was significantly reduced [8]. Interestingly, a previous research has demonstrated that the addition of fetal bovine exosomes declined contents of macrophage TNF-α and IL-6 [27]. A study has revealed that purified exosomes suppressed production of IL-1β in lipopolysaccharide/nigericin-stimulated macrophages [28]. Furthermore, Nong et al. have suggested that inflammatory markers, such as TNF-α and IL-6, were dramatically decreased after administration of exosomes produced through human-induced pluripotent stem cell-derived MSCs [29]. There is a article finding that the SBP was markedly elevated in the group of women who later developed PE [30, 31]. It was displayed that PE patients were positively associated with SBP and diastolic blood pressures and proteinuria [32]. Also, a recent study has provided a proof that proteinuria heightened with advancing gestation in PE women [33]. A important finding was that rats from the PE group had increased TNF-α relative to the normal pregnant group [34]. Another study has verified that serum IL-6 and IL-1β were obviously elevated in women with PE in relation to controls [35]. The above findings suggested that PE patients usually showed high SBP, proteinuria and levels of inflammatory factors. Thus, it could be inferred from our results that the hucMSC-derived exosomal miR-18b-3p had a therapeutic effect on PE.
Conclusion
In conclusion, our study provides evidence that hucMSCs-derived exosomes upregulate miR-18b-3p, which targets LEP to suppress the contents of inflammatory factors and reduce cell apoptosis rate in PE rat placenta tissues, thereby inhibiting the occurrence of PE. Thus, exosomal miR-18b-3p may be a potential candidate for treatment of PE via targeting LEP. This research identified the role of hucMSC-derived exosomal miR-18b-3p targeting LEP during PE development for the first time, which provided a novel insight for PE treatment. However, due to the limitation of known researches, the study needs to be monitored rigorously and reported appropriately in the future clinical trials.
数据和材料的可用性
Not applicable.
纳米材料