

多壁碳纳米管触发番茄侧根形成的新机制:一氧化氮的参与
摘要
大量研究表明,多壁碳纳米管(MWCNTs)对植物有毒。然而,MWCNT 是否或如何影响侧根 (LR) 的形成,这是根系统对各种环境因素的适应性的重要组成部分,仍然存在争议。在本报告中,我们发现多壁碳纳米管可以进入番茄幼苗根部。使用 MWCNTs 以近似剂量依赖性的方式促进番茄 LR 的形成。内源性一氧化氮 (NO) 的产生是由 MWCNTs 触发的,通过 Greiss 试剂法、电子顺磁共振 (EPR) 和激光扫描共聚焦显微镜 (LSCM) 以及 NO 的清除剂证实。 MWCNTs 和 NO 在 LR 发育的诱导过程中存在因果关系,因为 MWCNT 触发的 NO 合成和 LR 形成明显被内源性 NO 及其清除剂去除所阻断。 NO产生酶硝酸还原酶(NR)的活性响应于MWCNT而增加。钨酸盐对 NR 的抑制不仅会损害 NO 的产生,还会消除由 MWCNT 引发的 LR 形成。 N 的添加 G -硝基-l-精氨酸甲酯 (l-NAME) 是哺乳动物一氧化氮合酶 (NOS) 样酶的抑制剂,未能影响 LR 的形成。总的来说,我们提出 NO 可能作为下游信号分子参与 MWCNT 控制 LR 发展,至少部分通过 NR。
介绍
碳纳米管有许多生物学和生物医学应用 [1, 2]。由于独特的容易穿透细胞膜的能力,碳纳米管的生物安全性一直是一个争论的话题[3, 4]。同时,由于碳纳米管的生产和使用迅速增长,因此表征其对人类和哺乳动物以及最近在植物中的细胞毒性的详细机制变得很重要 [3,4,5,6,7,8,9]。众所周知,植物及其群落对人类和环境非常重要,而且它们也有暴露于碳纳米管的风险,因为通过施用生物固体肥料在土壤中积累 [6, 10, 11]。作为碳纳米管的重要成员,由多层石墨烯卷成的多壁碳纳米管(MWCNTs)的毒性已被广泛研究。哺乳动物研究表明,接触多壁碳纳米管和单壁碳纳米管会在人角质形成细胞和 A549 细胞中诱导氧化损伤和 NF-κB 活化 [9, 12]。多壁碳纳米管和单壁碳纳米管可以与质膜融合,从而通过脂质过氧化和氧化应激引起细胞损伤 [9, 11, 13, 14]。在人脐静脉内皮细胞中观察到由多壁碳纳米管引发的细胞毒性和氧化应激,以及适度的炎症反应 [15]。先前的研究表明,红菠菜中 MWCNTs 的主要毒性主要来自活性氧 (ROS) 的过度产生,并且可以通过补充抗坏血酸来逆转毒性作用 [7]。从这个意义上说,MWCNTs被认为是一种新的对动物或植物生物体的胁迫因子。
侧根 (LR) 的形成是根结构的重要决定因素,已被认为是对各种压力的适应性反应的指标 [16]。在高等植物中,LR 的形成受植物激素和各种环境因素的影响,包括水的可用性、养分和非生物胁迫,如缺氧和重金属胁迫 [17,18,19]。同时,大量证据证实,LR的形成不仅作为物理支持,而且有助于植物生长发育对水分和养分的吸收[19,20,21]。不同的环境线索可以触发几种特定的应激诱导形态发生反应 (SIMR) 表型,包括促进 LR 形成和抑制根伸长 [17]。LR 形成的调节也受到植物激素(如生长素、并且提出了响应生长素的细胞周期调节基因的激活 [19, 22]。同时,在黄瓜、番茄、大豆和油菜籽植物中证实了一些小分子参与生长素触发的根器官发生 [23,24,25,26,27]。这些小分子包括过氧化氢 (H2O2)、一氧化氮 (NO)、一氧化碳 (CO) 和氢气 (H2)。
其中,NO 是一种自由基气体,已被证明在植物中具有多种生理功能 [28, 29]。除了增强植物对逆境的适应能力外,NO 的功能还包括促进根毛发育、不定生根和侧根形成 [30,31,32,33],尽管在这些植物中 NO 生物合成的酶源上述过程仍然难以捉摸。在动物中,从 L-精氨酸合成 NO 是由含血红素的酶一氧化氮合酶 (NOS) [34] 催化的。尽管尚未在植物中鉴定出编码 NOS 酶的基因,但广泛检测到哺乳动物 NOS 样活性 [35, 36],以及哺乳动物 NOS 的抑制剂,如 N G -硝基-l-精氨酸甲酯盐酸盐 (l-NAME),可以抑制植物中 NO 的生成 [25, 33, 36,37,38,39]。重要的是,充足的遗传证据表明,NO 可以由硝酸还原酶 (NR) 产生,这是一种众所周知的负责植物氮代谢的酶 [28]。 NR 介导的 NO 产生参与气孔关闭和冷驯化已在遗传上得到证实 [37, 38]。我们之前的研究表明,NR依赖的NO合成参与了生长素诱导的氢气介导的侧根形成[39]。
到目前为止,在补充纳米材料时,在各种植物物种中分别报道了不同的 LR 形成、促进或抑制反应,包括 MWCNTs [40,41,42,43]、金纳米粒子(Au NP, [44])、氧化锌纳米粒子(ZnO NP [45, 46];)、二氧化钛纳米粒子(TiO2 NP [46];)和氧化石墨烯(GO [47,48,49];)(表 1),目前还没有研究提供明确的证明 NO 在上述反应中的作用。在本研究中,通过 Greiss 试剂法、激光扫描共聚焦显微镜 (LSCM) 和电子顺磁共振 (EPR) 分析检测内源性 NO,表明经 MWCNT 处理的番茄幼苗中 NO 水平增加。之后,观察到LR形成。我们通过使用抑制 NR 和哺乳动物样 NOS 活性的 NO 清除剂和拮抗剂来操纵内源性 NO 水平,进一步研究 NO 参与由 MWCNT 触发的 LR 形成。进一步的实验表明,NR 依赖性 NO 可能(至少部分地)是响应 MWCNT 形成 LR 所必需的。这项工作为了解纳米材料在植物中的生物学效应打开了一扇新窗口。
材料和方法
化学品
除非另有说明,所有其他化学品均从 Sigma-Aldrich(美国密苏里州圣路易斯)获得。从 Sigma-Aldrich 购买的 MWCNT 的特征如前所述 [50]。 MWCNTs的外径、内径和长度分别为6-12 nm、2.5-5 nm和1-9 μm。得到的匀浆胶体悬液经超声处理后灭菌备用。
其他碳纳米粒子购自南京XFNANO材料科技有限公司,包括单壁碳纳米管(SWCNTs,XFS22;纯度> 95%,直径1-2 nm,长度5-30 μm,比表面积> 1075 m 2 /g),石墨烯(XF001W;纯度~ 99%,直径0.5-5 μm,厚度~ 0.8 nm,单层比~ 80%,BET表面积500~1000 m 2 /G;电阻率≦ 0.30 Ω.cm),活性炭(AC,XFP06;纯度> 95%,粒径5 ± 1 μm,孔体积1-1.2 cm 3 /g,孔径2.0-2.2 nm,比表面积~ 1500-1700 m 2 /g).
此外,硝普钠 (SNP) 用作释放 NO 的化合物 [30,31,32,33]。 2-(4-Carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide 钾盐 (cPTIO) 被认为是 NO 的清除剂 [51,52,53,54]。钨酸盐(Tg;NR [28, 33, 37, 55,56,57] 的抑制剂;)和 N G -硝基-L-精氨酸甲酯盐酸盐(名称;哺乳动物 NOS 样酶的抑制剂 [25, 33, 36,37,38,39];)也被应用。本研究在中试实验中测定了上述化学物质的浓度,从中观察到显着的反应。
植物材料和生长条件以及LR形成的测定
番茄 (Solanum lycopersicum L.) 种子“江苏 14”由中国江苏省南京市江苏农业研究所友情提供。选择相同大小的种子在蒸馏水中在 25 ± 1 °C 下在黑暗中发芽 3 天。然后将选定的具有 2-3 mm 胚根的相同幼苗转移到 6 mL 处理溶液中,该溶液含有指定浓度的 MWCNT、200 nM 1-萘基乙酸(NAA;一种众所周知的生长素)、0.1 mM SNP、0.2 mM cPTIO、20 μM 钨酸盐 (Tg)、0.2 mM NAME 和其他碳纳米粒子,包括 5 mg/mL 单壁碳纳米管 (SWCNT)、石墨烯和活性炭 (AC),在指定的时间点单独或组合使用。幼苗在光照培养箱(25 ± 1 °C)中生长,光照强度为200 μmol m -2 s −1 14/10 h(明/暗)光周期。
处理后拍照,然后使用Image J软件(http://rsb.info.nih.gov/ij/)确定每株幼苗出现侧根的数量和长度(> 1 mm)[39 , 58]。如前所述,仅使用侧根诱导段进行后续分析。
通过透射电子显微镜成像 MWCNT 分布
使用透射电子显微镜(TEM;JEOL,JEM-200CX,东京,日本)表征 MWCNT 在番茄幼苗根中的分布。 TEM 分析的样品制备按照之前的协议[59]。
激光扫描共聚焦显微镜对内源性 NO 成像
NO 成像是通过使用相当特异性的 NO 荧光探针 4-amino-5-methylamino-2',7'-difluorofluorescein diacetate (DAF-FM DA) 进行的。彻底清洗探针后,使用 Zeiss LSM 710 共聚焦显微镜(Carl Zeiss,Oberkochen,德国,激发波长为 488 nm,发射波长为 500-530 nm 进行 NO 分析)获得图像。在我们的实验中,每个处理随机选择和测量 20 个个体样本。照片代表相同的结果。
Griess 试剂测定确定无内容
根据之前描述的方法 [50],NO 含量是用 Griess 试剂测定法测定的。重要的是,为了避免植物中高浓度硝酸盐和亚硝酸盐含量造成的干扰,将在 200 μM cPTIO(NO 清除剂)中预孵育 30 分钟的相同样品作为空白样品。加入Griess试剂30 min后,在540 nm处记录吸光度,与NaNO2标准曲线比较测定NO含量。
用电子顺磁共振 (EPR) 测定 NO
根据我们之前的方法 [39, 55, 60],使用电子顺磁共振 (EPR) 测定 NO 水平。有机溶剂层用于在 Bruker A300 光谱仪(Bruker Instrument,Karlsruhe,Germany)上在以下条件下测定 NO:室温;微波频率,9.85 GHz;微波功率,63.49 mW;和调制频率,100.00 kHz。
硝酸还原酶 (NR) 活性的测定
根据先前的方法[57],在540 nm处通过分光光度法检测NR活性。通过加入 1 mL 1% (w/v) 次磺酰胺的 3 M HCl 溶液以及 1 mL 0.02% (v/v) 正-(1-萘基)-乙二胺,在 540 nm 下分光光度法测定产生的亚硝酸盐。
统计分析
如果指出,结果表示为三个独立实验的平均值 ± SE,每个实验具有三个生物学重复。使用SPSS Statistics 17.0软件进行统计分析。对于统计分析,邓肯多重检验 (p 选择<0.05)。
结果
MWCNTs不仅进入根细胞,还促进LR形成
LR 的形成是根系统架构的主要决定因素。为了研究 MWCNTs 对 LR 形成的影响,将 3 天大的番茄幼苗与一系列浓度的 MWCNTs(0.05、0.5、5 和 50 mg/mL)孵育 3 天。 1-萘乙酸(NAA)的应用被视为阳性对照。在我们的实验中,LR 数量和长度都被确定为 LR 形成的两个参数。如图 1 所示,与对照样品相比,外源性 MWCNTs 以近似剂量依赖性的方式显着诱导 LR 形成,在 5 和 50 mg/mL 时效果最大。当施用 200 nM NAA 时,观察到类似的诱导反应。考虑到MWCNTs的成本和LR形成的诱导反应,5 mg/mL MWCNTs用于以下实验。
<图片>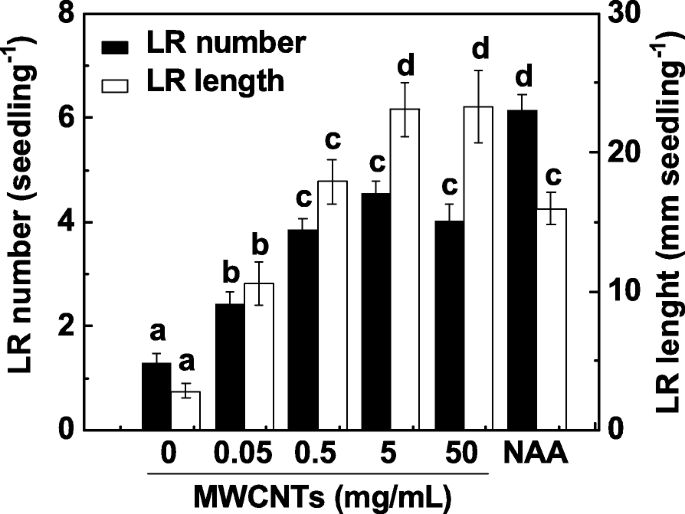
MWCNT 诱导的番茄 LR 形成大致呈剂量依赖性。三天大的番茄幼苗分别用 200 nM NAA 和指定浓度的 MWCNTs 处理。然后在处理 3 天后确定每株幼苗出现的侧根 (> 1 mm) 的数量和长度。 30株(10 × 3)植物,3个生物学重复,实验进行3次。数据是平均值 ± SE。在每组实验中,由相同字母表示的条形在 p 上没有显着差异 <0.05 根据邓肯多重检验的水平
为了验证 MWCNTs 在诱导 LR 形成中的特定功能,我们进一步研究了 MWCNTs 的其他同素异形体是否也具有这种诱导作用。如图 2a 所示,所有这些碳纳米材料都对枝条生长表现出毒性作用(数据未显示)。有趣的是,与不含化学物质的对照植物相比,MWCNTs、单壁碳纳米管(SWCNTs)、石墨烯和相同浓度(5 mg/mL)的活性炭的应用可能会导致LR数量和长度的增加(图2b)。在这些化学物质中,在多壁碳纳米管培养的番茄幼苗中发现了最大的诱导反应。
<图片>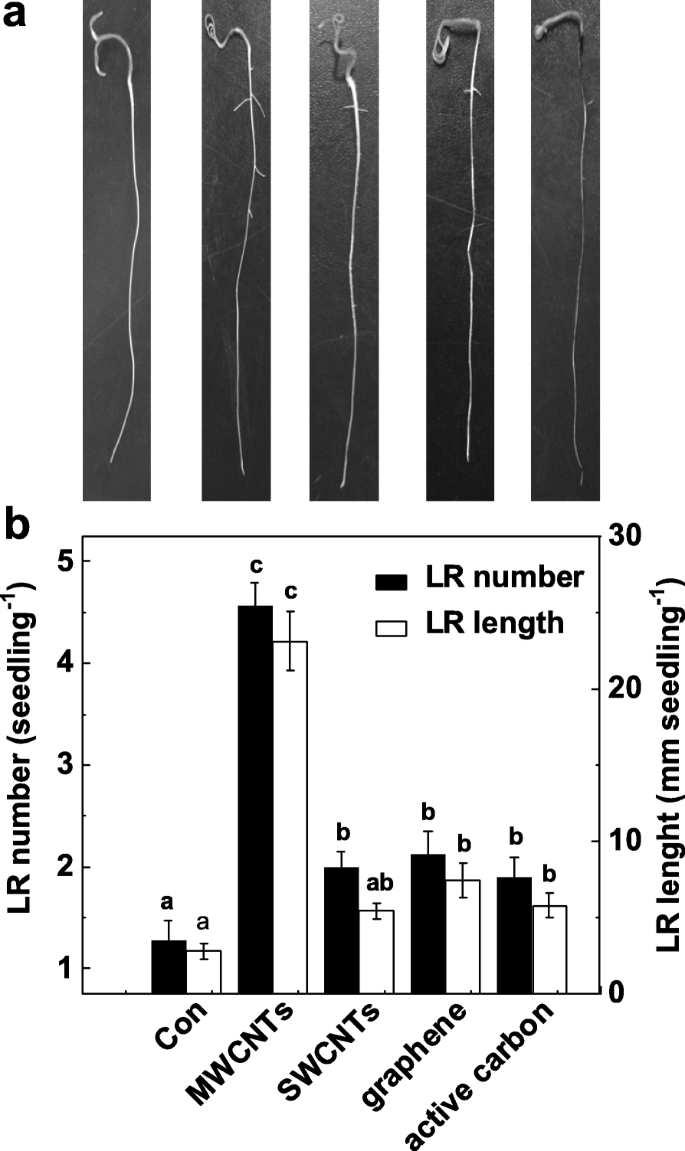
响应不同碳纳米粒子的侧根形成变化。将三天大的番茄幼苗分别用蒸馏水 (Con)、5 mg/mL MWCNT、单壁碳纳米管 (SWCNT)、石墨烯和活性炭 (AC) 再处理 3 天。 一 然后拍摄了代表性照片。 b 然后还确定了每株幼苗出现的侧根(> 1 mm)的数量和长度。比例尺 =50 mm。 30株(10 × 3)植物,3个生物学重复,实验进行3次。数据是平均值 ± SE。在每组实验中,由相同字母表示的条形在 p 上没有显着差异 <0.05 根据邓肯多重检验的水平
借助透射电子显微镜 (TEM),可以轻松评估 MWCNT 的分布。图 3 显示的结果表明,外源施用 MWCNTs 时,番茄幼苗可以吸收,并且发现 MWCNTs 分布在根细胞中。这个结果可以理解,因为幼苗根系直接在含有多壁碳纳米管的液体溶液中培养。
<图片>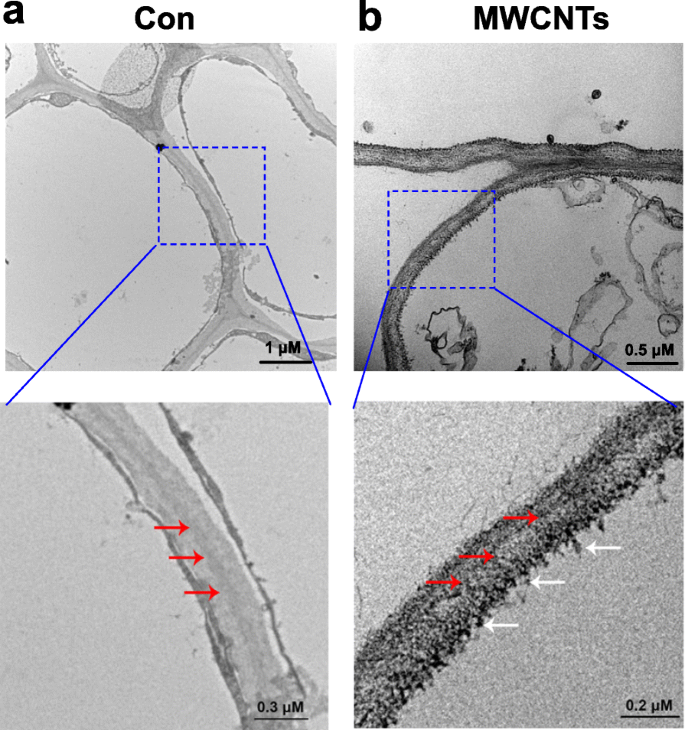
MWCNTs 在番茄根中的分布。用蒸馏水处理的 3 天龄番茄幼苗的 TEM 图像 (Con; a ) 或 5 mg/mL MWCNTs (b ) 1 天。红色箭头表示细胞壁,白色箭头表示多壁碳纳米管
MWCNT 诱导的 NO 合成和随后的 LR 形成对 cPTIO(NO 的清除剂)敏感
为了研究 NO 是否也参与 MWCNT 诱导的 LR 形成,通过使用 NO 释放化合物和清除剂操纵内源性 NO 水平来评估 NO 在 MWCNT 引起的 LR 形成中的功能。与之前的结果 [31] 类似,硝普钠 (SNP) 的施用可导致 LR 形成的诱导,并且当 SNP 和 MWCNTs 一起应用时观察到加性反应(图 4)。当加入 2-(4-carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide 钾盐(cPTIO;NO 的清除剂)时,MWCNTs 对 LR 形成的促进反应显着受损。与不含化学物质的对照相比,单独使用 cPTIO 可以抑制 LR 发育,表明内源 NO 在根器官发生中的重要作用。
<图片>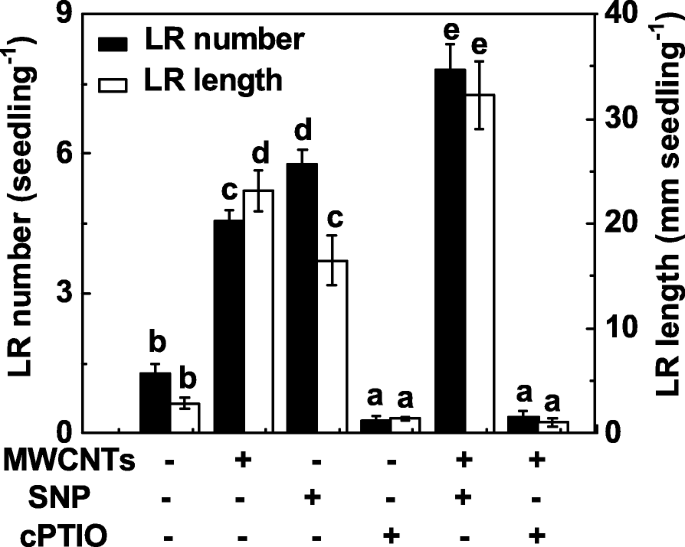
MWCNT 诱导的 LR 形成对用 cPTIO(其清除剂)去除内源性 NO 很敏感。用蒸馏水、5 mg/mL MWCNT、0.1 mM SNP、0.2 mM cPTIO,单独或组合处理三天大的番茄幼苗3 天。然后,然后确定每株幼苗出现的侧根(> 1 mm)的数量和长度。 30株(10 × 3)植物,3个生物学重复,实验进行3次。数据是平均值 ± SE。在每组实验中,由相同字母表示的条形在 p 上没有显着差异 <0.05 根据邓肯多重检验的水平
为了进一步评估内源性 NO 在 MWCNT 反应中的重要作用,首先用 Greiss 试剂法检测了体内 NO 产生的时间过程。在上述测定过程中,以cPTIO预处理的相同滤液为准确结果的空白。据观察,番茄幼苗根部的 NO 产生显着增加,直到 MWCNT 处理后 24 h,然后恢复到初始水平(48 h;图 5a)。 24 h由MWCNTs触发的高于最大水平的内源性NO被NO清除剂cPTIO明显消除,表明NO的特殊作用。
<图片>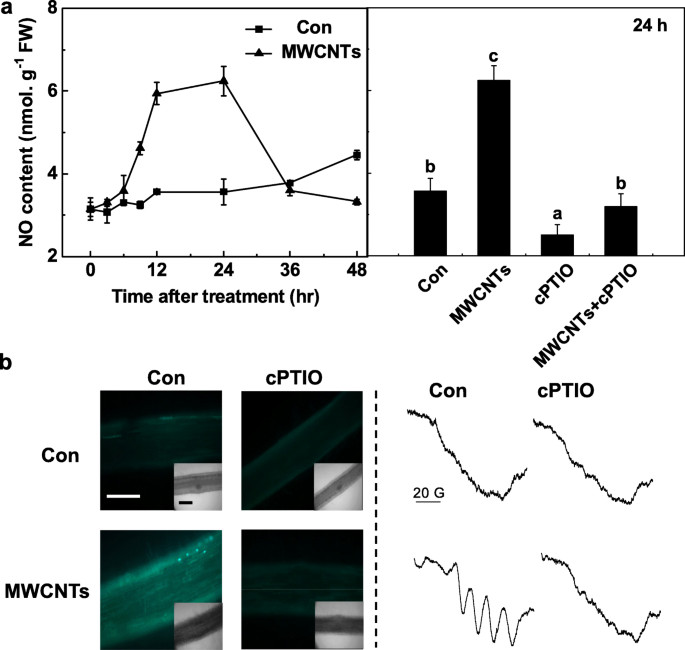
MWCNT 诱导的 NO 产生被 NO 清除剂 cPTIO 阻断。三天大的番茄幼苗分别用蒸馏水和 5 mg/mL MWCNTs 处理,分别含有或不含有 0.2 mM cPTIO。 一 使用 Greiss 试剂方法测定的 NR 活性(左)和 NO 产生(右)的变化。 b 治疗24 h后,通过LSCM(左)和EPR(右)分析NO信号。比例尺 =0.1 mm。数据是平均值 ± SE。由相同字母表示的条形在 p 处没有显着差异 <0.05 水平,根据 Ducan 的多重检验
为了确认上述结果,LSCM 和 ESR 都被采用。首先,番茄幼苗根部内源性 NO 水平的变化通过使用细胞渗透性、相当 NO 特异性的荧光探针 DAF-FM DA 标记 NO 并用 LSCM 成像来监测。与之前的结果(图 5a)类似,在 cPTIO 的存在下,由 MWCNT 触发的 DAF-FM 依赖性荧光强度的增加被大大消除(图 5b)。这些结果表明,DAF-FM触发的荧光与番茄幼苗根部内源性NO水平有关。
EPR 光谱证实了 MWCNT 诱导的 NO 产生。正如预期的那样,用 MWCNTs 处理 24 h 的幼苗根部呈现出典型的 NO 复合物的超精细结构三重态。然而,cPTIO 的添加消除了上述信号,表明 MWCNT 暴露确实导致了强烈的 NO 产生(图 5b)。总的来说,这些数据表明在番茄幼苗中MWCNT触发的LR形成可能需要NO合成。
NR 可能负责 MWCNT 诱导的 NO 产生和随后的 LR 形成
由于NR和类哺乳动物NOS是植物中与NO合成相关的两种主要酶,因此在随后的实验中同时应用了钨酸盐(一种NR抑制剂)和NAME(一种哺乳动物NOS抑制剂)。在这里,钨酸盐处理显着抑制了 MWCNT 处理的番茄幼苗根中 LR 形成的促进(图 6)。相比之下,添加 NAME 并没有强烈抑制由 MWCNT 引发的 LR 形成的诱导,表明哺乳动物样 NOS 可能不是负责由 MWCNT 引发的 NO 产生的目标 NO 合成酶。还观察到,当分别施用钨酸盐或 NAME 时,番茄幼苗中 LR 的形成略有但没有显着降低。
<图片>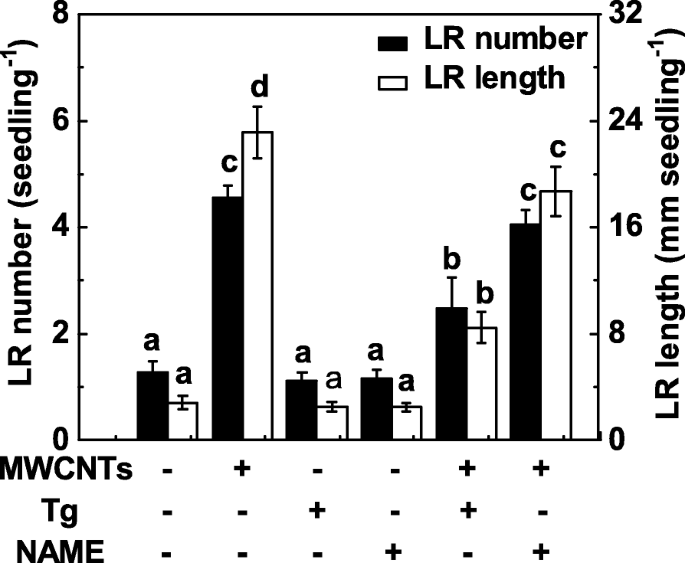
响应于 MWCNTs 和两种 NO 合成抑制剂的 LR 形成变化。用 5 mg/mL MWCNT、20 μM 钨酸盐 (Tg)、0.2 mM NAME 单独或组合处理三天大的番茄幼苗 3 天。然后,然后确定每株幼苗出现的侧根(> 1 mm)的数量和长度。 30株(10 × 3)植物,3个生物学重复,实验进行3次。数据是平均值 ± SE。在每组实验中,由相同字母表示的条形在 p 上没有显着差异 <0.05 根据邓肯多重检验的水平
NR 在 MWCNT 触发的 LR 形成中的作用通过监测 NO 产生以响应应用的 MWCNTs 有或没有钨酸盐来进一步检查。与内源性 NO 产生的变化(图 5a)相比,NR 活性的时程分析显示出类似的趋势(图 7a),也在用 MWCNTs 处理后 24 h 达到峰值。这些结果表明 MWCNT 诱导的 NO 产量增加可能主要是由于 NR 的活性增强。一致地,通过使用 Greiss 试剂方法(图 7b)、LSCM 和 EPR(附加文件 1:图 S1)证实了钨酸盐对 NR 依赖性 NO 产生的抑制。
<图片>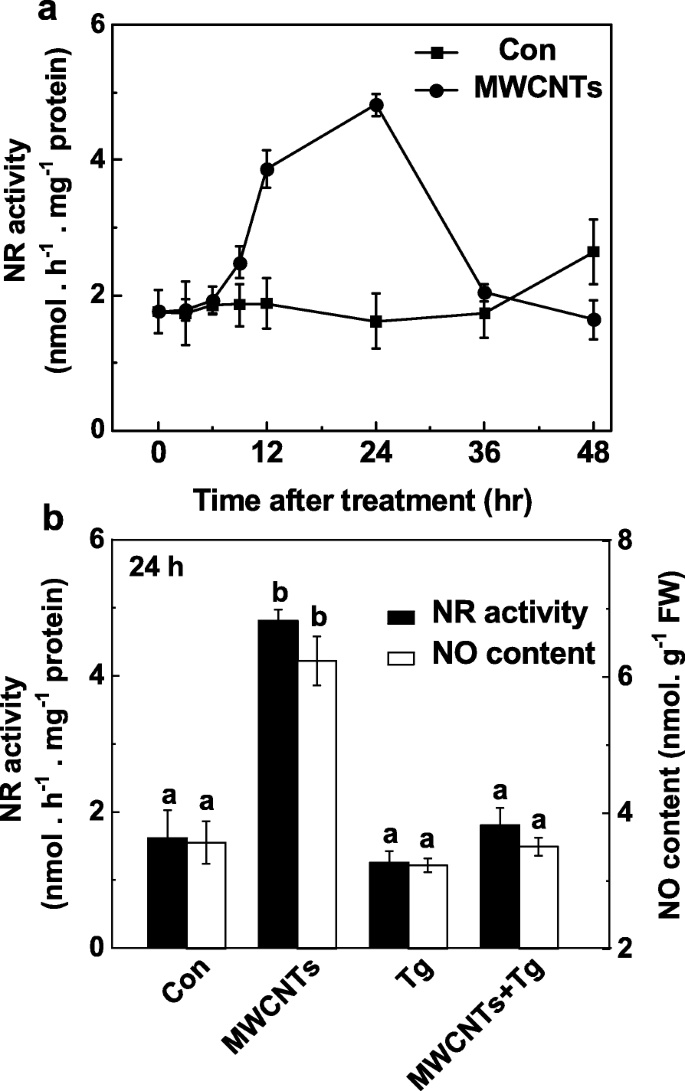
MWCNT 诱导的 NO 产生被 NR 抑制剂钨酸盐阻断。三天大的番茄幼苗用蒸馏水和 5 mg/mL MWCNTs 处理,有或没有 20 μM 钨酸盐 (Tg)。 NR 活动的变化 (a ) 和 NO 产生 (b ) 使用 Greiss 试剂法测定。数据是平均值 ± SE。由相同字母表示的条形在 p 处没有显着差异 <0.05 水平,根据 Ducan 的多重检验
讨论
植物毒性是了解纳米粒子潜在环境影响的重要考虑因素 [4, 7, 61,62,63]。大量证据表明,多壁碳纳米管对植物具有毒性,包括诱导氧化损伤、抑制种子萌发、根系生长和发育 [11, 63, 64]。然而,作为 SIMR 的一种表型,通过侧根形成的根分支是根系统对各种环境因素的适应性的重要组成部分 [17]。在这项工作中,我们整合了生物学、药理学和生化分析,以显示 NR 介导的 NO 产生参与 MWCNT 诱导的 LR 形成,至少部分是在我们的实验条件下。此外,一氧化氮在多壁碳纳米管刺激的根器官发生中的功能强调了这种第二信使在植物发育过程和逆境适应中的核心作用[29,30,31,32,33,37,38]。
首先,我们证实 5 mg/mL MWCNTs(OD 6-12 nm)可以进入根组织(图 3)。之后,观察到番茄 LR 形成的诱导(图 1),模拟 NAA 和 SNP 的诱导作用(图 4),这是一种众所周知的 NO 释放化合物 [30, 31]。在用多壁碳纳米管(OD 6–13 nm、约 9.5 nm 和 30–40 nm,分别为 OD 6–13 nm)攻击时,在树脂树 [41]、生菜 [42] 和拟南芥 [43] 中发现了类似的诱导反应。例如,应用原始多壁碳纳米管(p-MWCNT)或羧基功能化多壁碳纳米管(c-MWCNT)(平均直径 9.5 nm)可以促进 LR 在生菜幼苗中的发育 [42]。相比之下,当受到多壁碳纳米管 (OD 20–70 nm [40];) 时,大豆植物中同时发现了初级根的抑制甚至 LR 的形成。通过与多壁碳纳米管外径数据(表1)的比较,我们认为多壁碳纳米管对LR形成的影响随其直径的不同而变化,显示出直径越小促进,直径越大抑制。当然,应该仔细研究相关机制。类似的现象在植物耐盐性中得到证实 [50]。结合上述结果,进一步推论纳米材料的功能可能因物种而异,反之亦然,不同类型的纳米材料可能会引起不同的生物学效应。然而,其他影响因素,如不同剂量的多壁碳纳米管 [48] 甚至植物生长条件等,也不能轻易排除。
与其他纳米材料(包括具有相同浓度的 SWCNT、石墨烯和 AC)相比,在 MWCNT 中观察到 LR 形成的最大诱导甚至对枝条生长的毒性作用(图 2)。这些可能与 MWCNTs 的特殊物理特性有关,这是一种具有高导电性、大比表面积、高纵横比和显着热稳定性的纳米材料 [65]。纳米材料的毒性作用在黄瓜、卷心菜、胡萝卜、洋葱等中已有广泛报道[66, 67]。
大量证据表明,NO 作为信号分子,可以调节从环境适应到发育的广泛植物过程,后者包括种子萌发和根器官发生 [29, 68,69,70,71,72,73 ]。我们随后的实验表明 NO 可能参与了 MWCNT 诱导的 LR 形成。虽然已经应用了几种对植物细胞中 NO 产生进行成像的方法,但每种方法都可能存在缺点,包括缺乏灵敏度和 NO 非依赖性分子的干扰 [74]。因此,在我们的实验条件下应用了三种负责 NO 成像和测定的方法,包括 Greiss 试剂法、LSCM 和 EPR,以及 cPTIO(一种 NO 清除剂)的应用。通过使用三种方法,我们观察到由 MWCNTs 在番茄幼苗中诱导的内源性 NO 产生增加被 cPTIO 消除(图 5),一种 NO 清除剂 [30,31,32]。重要的是,该过程与 MWCNT 诱导的 LR 发育的生物学反应相关,当 cPTIO 同时应用时,LR 发育被严重阻断(图 4)。
对这些反应和外源性应用 MWCNT 诱导的 NO 的潜在来源的进一步评估表明,NO 的产生和随后的 LR 形成可归因于 NR 活性。在植物中,NO 的产生主要来自 NR 和哺乳动物 NOS 样蛋白 [28]。然而,植物 NOS 基因仍未确定 [35, 75, 76],尽管一些使用哺乳动物 NOS 酶抑制剂的实验提供了一些证据,证明 NO 生产中 L-精氨酸依赖途径 [36, 76]。 NR 被证实是植物中最重要的 NO 来源 [28]。先前的研究表明,依赖于 NR 的 NO 产生作为硝酸盐相关信号,参与根结构的调节 [32, 33]。此外,依赖于 NR 的 NO 产生与冷驯化 [38]、耐盐性 [50] 和脱落酸诱导的气孔关闭 [77] 密切相关。我们的结果进一步表明,钨酸盐(NR 的抑制剂)明显损害了 MWCNT 诱导的 LR 形成,尤其是在 LR 长度方面(图 6)。相比之下,当使用 l-NAME(哺乳动物 NOS 抑制剂)时,LR 长度仅略有减少,并且 LR 数量没有显着差异。一致地,生化分析表明 MWCNTs 显着增加了 NR 活性(图 7a),与 NO 产生的变化平行(图 5a)。钨酸盐可以完全阻止上述反应(图 7a,附加文件 1:图 S1)。因此我们推断,由多壁碳纳米管诱导的内源性 NO 产生的增加主要归因于 NR 途径。 Certainly, further genetic evidence should be investigated.
Conclusion
In summary, we provide evidence to show that MWCNT-induced NO production via NR might be required for tomato lateral root formation and this was summarized in Fig. 8. Importantly, above findings provide insights into the intricate molecular mechanism of MWCNTs functions in plants.

Schematic representation of the proposed MWCNT-induced tomato lateral root formation mainly via NR-dependent NO production. The role of mammalian NOS-like enzyme was preliminarily ruled out
数据和材料的可用性
All data are fully available without restriction.
缩写
- CO:
-
Carbon monoxide
- cPTIO:
-
2-(4-Carboxyphenyl)-4,4,5,5-tetramethylimidazoline-1-oxyl-3-oxide potassium salt
- DAF-FM DA:
-
4-Amino-5-methylamino-2′,7′-difluorofluorescein diacetate
- EPR:
-
Electron paramagnetic resonance
- 开始:
-
氧化石墨烯
- H2:
-
Hydrogen gas
- H2O2:
-
Hydrogen peroxide
- l-NAME:
-
N G -Nitro-l-arginine methyl ester
- LR:
-
Lateral root
- LSCM:
-
Laser scanning confocal microscopy
- MWCNTs:
-
Multi-walled carbon nanotubes
- NAA:
-
1-Naphthylacetic acid
- NO:
-
Nitric oxide
- NOS:
-
Nitric oxide synthase
- NR:
-
Nitrate reductase
- ROS:
-
Reactive oxygen species
- SIMR:
-
Stress-induced morphogenic response
- SNP:
-
Sodium nitroprusside
- SWCNTs:
-
Single-walled carbon nanotubes
- TEM:
-
透射电子显微镜
- Tg:
-
Tungstate
纳米材料