

细菌微室壳蛋白对环境变化的自组装稳定性和变异性
摘要
细菌微区室 (BMC) 是蛋白质自组装细胞器,在原核生物界中广泛存在。通过使用多面体壳分割关键代谢酶和途径,BMC 在碳同化、发病机制和微生物生态学中发挥着重要作用。 BMC 壳由多个蛋白质同源物组成,它们自组装形成定义的结构。人们对工程化 BMC 以开发新的纳米生物反应器和分子支架产生了极大的兴趣。在这里,我们使用高速原子力显微镜 (HS-AFM) 报告了在不同 pH 和盐条件下 BMC 壳蛋白的形成和自组装动力学的定量表征。我们表明,400 mM 的盐浓度容易导致由壳六聚体形成的更大的单层壳斑块,并且在中性 pH 值下观察到更高的六聚体自组装动态速率。我们还可视化了壳蛋白从六聚体组装到纤维状阵列的可变性。这项研究提高了我们对 BMC 蛋白质自组装响应微环境变化的稳定性和可变性的认识,这将为合成 BMC 结构的合理设计和构建提供信息,并具有重塑其自组装和结构稳健性的能力。它还提供了一个强大的工具箱,用于定量评估生物技术应用中基于 BMC 的纳米结构的自组装和形成。
介绍
细菌微区室 (BMC) 是蛋白质细胞器,在结构上类似于病毒衣壳,可分隔细菌的细胞质 [1]。它们在细菌门中广泛存在 [2],并允许细菌在真核生物中缺乏膜结合细胞器的情况下划分关键的代谢途径 [3, 4]。 BMCs 由包裹着管腔酶核心的半透性蛋白质外壳形成。外壳由三种结构蛋白成分组成,包括BMC-H(包含一个Pfam00936域)、BMC-T(包含两个Pfam00936域)和BMC-P(包含一个Pfam03319域)[5,6,7, 8,9]。壳的主要成分是 BMC-H,它表现为具有凸面和凹面的六聚体,并平铺壳面,凹面朝外 [10](图 1)。 BMC-P 形成五聚体,被提议覆盖二十面体形状的顶点,BMC-T 形成位于壳面的假六聚体,可能负责壳渗透性。
<图片>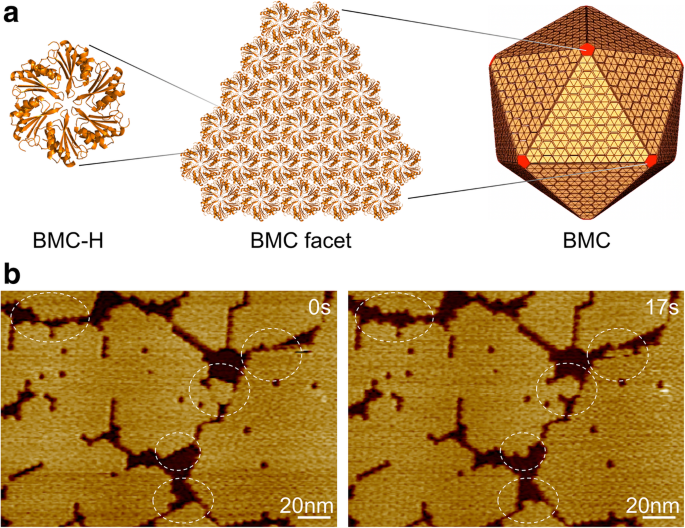
细菌微室、壳组织和自组装。 一 数百个 BMC 壳蛋白同系物自组装形成二十面体蛋白细胞器。黄色的 BMC-H 蛋白形成小平面,红色的 BMC-P 蛋白占据顶点。 b 由 Hoch_5815 BMC-H 六聚体组成的壳面的 AFM 形貌。使用 HS-AFM 在数秒内观察到动态事件(圆圈)
特定的蛋白质-蛋白质相互作用确保 BMC 蛋白质的自组装形成高度明确的结构,以实现其代谢功能。壳蛋白之间的横向相互作用被认为是决定二十面体壳自组装特性的主要因素 [10]。已经观察到 BMC-H 同系物可以形成各种形状,包括二维片 [11, 12]、纳米管 [13,14,15,16,17] 和细丝结构 [15,18,19,20] .
基于天然细胞器的自组装、选择性渗透和酶包封特性,BMCs 被认为是一种在生物工程中具有巨大潜力的理想系统,包括通过包裹代谢酶和生成具有新功能 [21,22,23,24,25,26]。然而,BMC 生物工程仍有一些关键问题有待解决,例如 BMC 结构的稳定性以及如何有效操纵和评估 BMC 蛋白质聚集体的自组装和形成。已经使用 X 射线晶体学、电子显微镜 (EM)、荧光显微镜和动态光散射 (DSL) [10, 11, 16, 22, 27,28] 对 BMC 壳和整个 BMC 的结构和组装进行了研究,29,30,31]。最近,我们利用高速原子力显微镜(HS-AFM)对BMC-H蛋白的动态自组装过程进行了首次可视化[12]。
在这项工作中,我们使用 HS-AFM 来监测 BMC-H 贴片在不同 pH 和离子条件下的结构动力学,这提供了对 BMC 壳蛋白组装调节的深入了解,并提供了一个强大的定量评估工具,在分子分辨率下, 关于 BMC 壳蛋白自组装的稳定性和可变性。
方法
样品准备
来自Haliangium ocraceum的纯化BMC-H蛋白(Hoch_5815) 由 Kerfeld 博士(劳伦斯伯克利国家实验室)友情提供。对于缓冲液交换,库存样品在 ~ 80 mg mL −1 在 Tris 缓冲液(50 mM Tris-HCl,pH 7.8,100 mM KCl,10 mM MgCl2)中稀释至 0.5 mg mL -1 在 AFM 成像之前使用所需的缓冲区(附加文件 1:图 S1)。对照缓冲液为 50-mM Tris-HCl (pH 7.8) 和 10 mM MgCl2。
原子力显微镜
所需的缓冲液用于云母上的样品吸收和 AFM 成像。在云母上吸收 5 分钟后,用所需的缓冲液冲洗 Hoch_5815 以去除固定的蛋白质,然后使用 AFM 成像(附加文件 1:图 S1)。 HS-AFM 图像是在 AC 模式的溶液中以 30 或 40 Hz 的频率使用配备有 ULTRA Speed 2.8 μm 扫描仪和超短悬臂 USC-0.3 MHz 探针(NanoWorld)的 JPK NanoWizard ULTRA 速度 AFM 捕获的。在AFM成像过程中施加~ 100 picoNewton的最小载荷力以减少蛋白质组装的干扰[12, 32,33,34,35,36]。
图像处理与分析
最初使用 JPK SPM 数据处理 (JPK) 进行图像分析。 HS-AFM 图像分析是使用 Image SXM (http://www.ImageSXM.org.uk) 上的自定义宏进行的,如前所述 [12]。为了分析 Hoch_5815 斑块的大小,以 30 Hz 扫描速率捕获的 512 × 512 像素的图像被展平以消除任何 XY 倾斜和 Z 阈值,然后进行二进制转换以显示蛋白质与非蛋白质。粒子分析用于计算这些二值图像中蛋白质的表面积。补丁被定义为由> 3 个像素(~ 2 nm)分隔的对象,以便识别单个补丁与相邻补丁。最初的测试表明,如果设置了更多的像素,相邻的补丁可以算作一个连续的补丁,而使用较小的像素数,补丁中各个六聚体之间的间隙可能会被误认为是补丁之间的边界。为了分析蛋白质动力学,分析了以 40Hz 扫描速度捕获的 256 × 256 像素的图像系列,给出了每帧大约 6.4 s 的时间分辨率。从系列中的先前图像中减去二值图像以显示不同的 AFM 图像。差异图像的粒子分析用于计算组装和分解蛋白质的面积。用于计算动态速率的公式如下:
$$ \mathrm{Rate}\ \mathrm{of}\ \mathrm{dynamic}\ \mathrm{events}\ (R)=\frac{\mathrm{Number}\ \mathrm{of}\ \mathrm{hexamers} \ \mathrm{已添加}\ \mathrm{or}\ \mathrm{已删除}\ \mathrm{in}\ \mathrm{a}\ \mathrm{s}\mathrm{eries}\ \mathrm{of}\ \mathrm {frame}\mathrm{s}\kern0.5em (N)}{\mathrm{Total}\ \mathrm{surface}\ \mathrm{area}\ \mathrm{of}\ \mathrm{protein}\ \mathrm{在}\ \mathrm{frame}\kern0.5em (A)\times \mathrm{time}\kern0.5em (T)}, $$其中 N 表示阈值差异图像中白色和黑色像素的总和除以对应于该比例的单个六聚体的像素数(附加文件 1:图 S3,图 S5)。数据表示为平均值 ± 标准偏差(SD)。使用多元方差分析或指定的双向方差分析进行统计分析。
结果
我们使用了来自粘杆菌 Haliangium ocraceum 的 BMC-H 蛋白 (Hoch_5815) , 在大肠杆菌中表达 并表征为具有六重对称性的六聚体 [12]。 Hoch_5815 六聚体可以在第二个时间尺度自组装形成单层片,这代表了二十面体 BMC 架构的基本结构组件(图 1a)。 HS-AFM 成像使我们能够可视化片断片的动态组装和组织灵活性(图 1b),并使用开发的成像分析定量估计 BMC-H 蛋白的斑块大小和动态速率(参见“方法”部分)。
对 pH 变化的响应
我们测量了补丁大小的变化,作为 Hoch_5815 自组装整体能力的指标。斑块大小随着 pH 从 3 升高到 10 而增加(图 2a;附加文件 1:图 S2、表 S1),表明高 pH 比低 pH 条件更有利于 Hoch_5815 蛋白质的自组装。这与 RmmH 蛋白的组装行为有些不同,后者被发现在 pH 6 时不溶,在 pH 8 时形成有序的纳米管阵列,并在 pH 10 时易于分解 [13]。此外,我们观察到 HOCH_5815 自组装的高度结构可变性(如图 2a 中的大 SD 所示,附加文件 1:图 S2)。
<图片>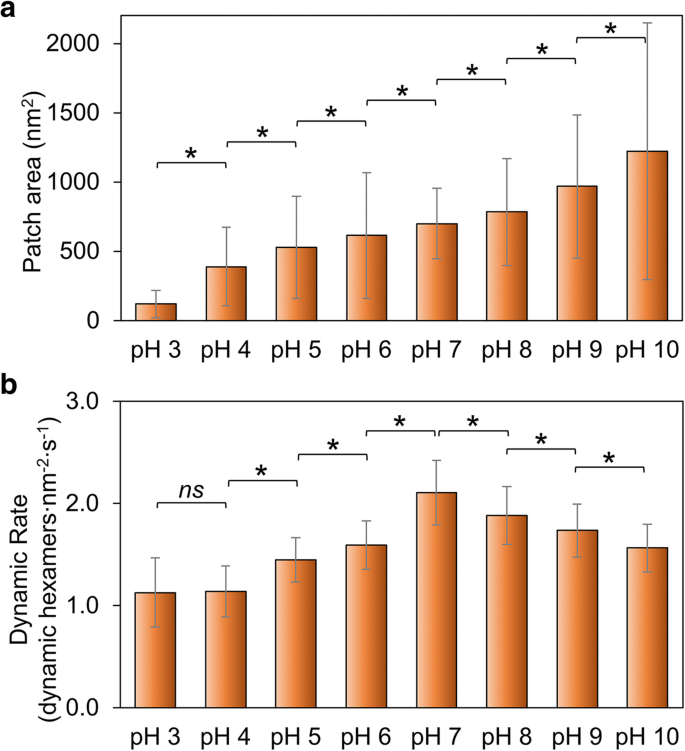
环境pH值对Hoch_5815自组装的影响。 一 原子力显微镜测定的 Hoch_5815 单个斑块的平均表面积 (n =50)(附加文件 1:图 S2)。 b 由 HS-AFM (n =50)。 *p <0.05, ns 不显着(多元方差分析)
对壳片中 Hoch_5815 蛋白质自组装的 AFM 成像表明,壳片的形成归因于六聚体的组装和拆卸的组合 [12]。我们进一步检查了不同 pH 值下 Hoch_5815 自组装动力学和动态事件的速率(附加文件 1:表 S2),以探索 Hoch_5815 蛋白质-蛋白质相互作用的稳定性。自组装动力学的速率在 pH 7 时最高,在酸性和碱性条件下均降低(图 2b;附加文件 1:图 S3)。特别是,它在酸性条件下迅速下降,特别是从 pH 7 到 pH 6,并在 pH 4 和 pH 3 之间出现相对恒定,如图 2b 所示。
pH 值可能对位于六聚体-六聚体界面的氨基酸残基的静电特性有很大影响。在酸性条件下观察到的动力学降低和壳斑块尺寸变小说明 Hoch_5815 具有降低的自组装能力。在碱性条件下观察到的动力学降低和壳斑块的更大尺寸表明六聚体-六聚体相互作用稳定,而 Hoch_5815 六聚体的动力学增加意味着在中性 pH 条件下六聚体-六聚体相互作用灵活。
对盐浓度变化的响应
我们还验证了缓冲液的盐浓度是否对 Hoch_5815 的组装有影响。在低浓度 (100–200 mM) MgCl2、CaCl2 和 KCl 下,Hoch_5815 蛋白形成的斑块比在较高浓度 (300–500 mM) 下组装的斑块相对较小(图 3a;附加文件 1:图 S4)。在 500 mM 时,我们观察到双层或多层 Hoch_5815 片材(附加文件 1:图 S4)。这些观察结果与先前的发现一致,即更高的离子强度可以促进 CcmK 形成更广泛和有序的 2D 晶体,CcmK 是用于碳同化的羧基体的壳蛋白 [37]。然而,当NaCl浓度从50增加到500 mM时,由RmmH形成的高度有序的纳米管被分解[13],表明介导壳六聚体形成平板和管状形状的潜在机制不同。
<图片>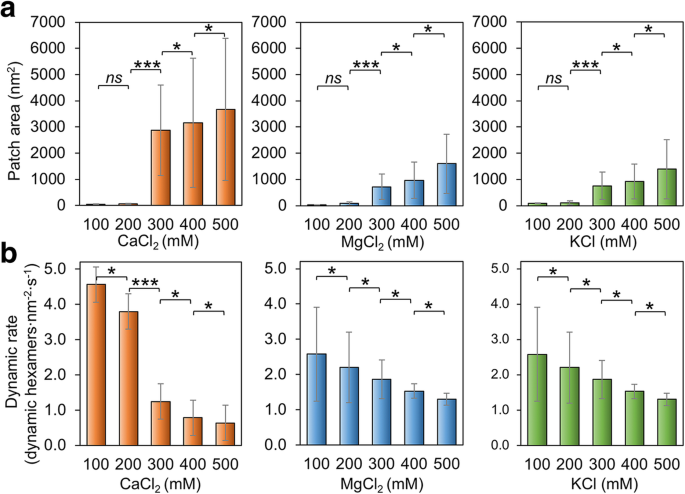
盐浓度对Hoch_5815自组装的影响。 一 在 100–500 mM CaCl2、MgCl2 和 KCl (n =50)。盐浓度的升高导致斑块尺寸增加。在 200 和 300 mM 之间观察到斑块面积的显着变化 (***p <0.001, *p <0.05, ns 不显着,双向方差分析)。 b 在 100–500 mM CaCl2、MgCl2 和 KCl (n =50)。盐浓度每变化 100 mM 都会导致动态事件发生率发生显着变化 (***p <0.001, *p <0.05, ns 不显着,双向方差分析)
此外,由MgCl2和KCl浓度变化引起的Hoch_5815自组装变化相对相似。相比之下,当 CaCl2 浓度从 200 增加到 300 mM 时,斑块大小的变化最为明显(增加了 3000 倍)(图 3a),这表明 Hoch_5815 自组装对 CaCl2 的敏感性高于对 CaCl2 的敏感性。 MgCl2 或 KCl。
Hoch_5815 自组装的动态速率也受缓冲盐浓度变化的影响。 MgCl2、CaCl2 或 KCl 浓度的增加可能导致 Hoch_5815 动态速率的下降(图 3b;附加文件 1:图 S5)。鉴于在较高盐浓度下观察到的斑块大小增加(图 3a),Hoch_5815 六聚体之间的横向相互作用似乎在高盐浓度下更稳定。 CaCl2 浓度的变化有更明显的响应,动态事件发生率在 200 和 300 mM 之间有显着变化(图 3b),而对 MgCl2 和 KCl 变化的响应相对相似,与补丁大小的变化(图 3a)。有趣的是,在 400 mM 的 MgCl2、CaCl2 或 KCl(附加文件 1:表 S2)下观察到组装事件与拆卸事件的最高比例。这导致在 400 mM 盐下形成大而稳定的单层 Hoch_5815 组件(附加文件 1:图 S4)。在 500 mM 下观察到的双层组件也很稳定,六聚体运动速率较低。
BMC-H 蛋白质组装的灵活性
通过将扫描力降低到 100 pN,我们最大限度地减少了 AFM 尖端扫描对 BMC 蛋白质组装的影响,并获得了单个六聚体的分子分辨率 AFM 图像(图 4)。组装和拆卸事件都可以在同一视图中看到,验证了 BMC 壳组装而不是尖端扫描伪影的动态特性 [12]。 HS-AFM 成像还揭示了 Hoch_5815 蛋白质聚集的可变性。在仅存在 10 mM MgCl2 的情况下在 pH 7.5 下对样品进行成像时,令人惊讶的是,我们偶尔会观察到纤维状结构的形成以及第二个时间尺度的 Hoch_5815 六聚体的分解(图 4a)。这些纤维状结构可以平行密集堆积,类似于由壳六聚体组装的纳米管束 [13,14,15,16]。然而,两根光纤之间的间距为 3.72 ± 0.31 nm (n =30),它们的平均高度为 2.46 ± 0.22 nm (n =30),小于由Hoch_5815六聚体形成的壳片(3.45 ± 0.16 nm,n =25)(图 4b-f)。这些纤维结构在成像过程中相当灵活和动态,可以显示不同尺寸的直线或盘绕结构。考虑到纤维结构与 Hoch_5815 六聚体的分解同时出现,并且与单层六聚体片相比它们的高度降低,我们推测这些纤维状结构是由从六聚体中分解出来的单个 Hoch_5815 肽形成的(图 4g) .在特定缓冲条件(例如低离子强度)中的底物吸收可能导致 Hoch_5815 肽的 α 螺旋侧附着到底物表面以及肽与相邻肽的线性结合,尽管假设内六聚体相互作用可能很强 [5]。壳蛋白聚集变异的详细机制仍有待进一步阐明。
<图片>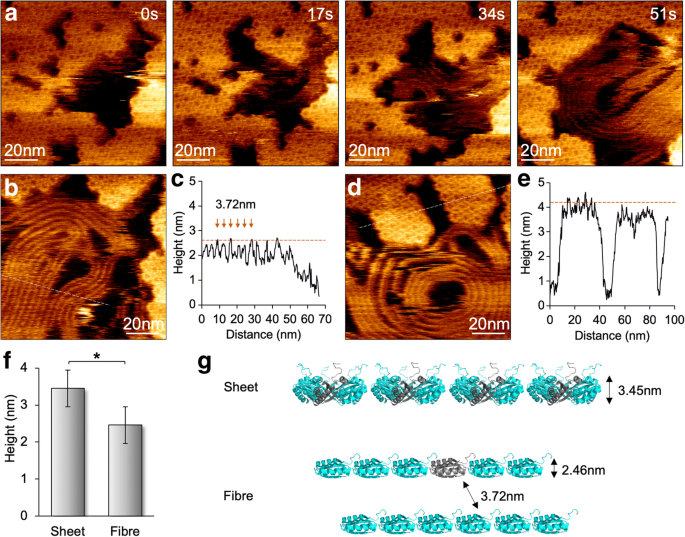
纤维结构的形成和动力学以及 HS-AFM 下的壳板组件。 一 由 Hoch_5815 六聚体组成的壳片在拆卸过程中出现的纤维状结构,如 AFM 图像系列所示。 b 纤维结构的 AFM 形貌图。 c 横截面分析(面板 b 中的虚线 ) 显示间距为 3.72 ± 0.31 nm (n =30) 相邻纤维结构之间,平均高度为 2.46 ± 0.22 nm (n =30)。 d 由 Hoch_5815 六聚体组成的壳贴片的 AFM 地形图。 e 横截面分析(面板 d 中的虚线 ) 表明 Hoch_5815 六聚体的平均高度为 3.45 ± 0.16 nm (n =25)。 f 与由 Hoch_5815 六聚体 (*p <0.05,双向方差分析)。 g 拟议的纤维状结构的组织和形成,代表一串Hoch_5815单体
讨论
BMC 包含数百种蛋白质,这些蛋白质自组装形成更高级的结构。 BMC 壳由众多蛋白质同源物组成,是研究蛋白质自组装和相互作用的理想系统。作为分析与生物系统生理作用高度相关的生物膜组织、蛋白质组装和物理相互作用的强大技术 [32, 35, 38, 39],AFM 已被用于可视化组织和自组装动力学BMC 壳蛋白以及 BMC 结构的结构和机械特征 [12, 30, 31, 40,41,42]。据我们所知,这项工作代表了第一次定量测定 BMC 壳蛋白在形成二维片以响应环境变化时使用 AFM 的自组装动力学。结果突出了 BMC-H 蛋白自组装的内在可变性和环境依赖性。与 EM 和 DSL 相比,AFM 在实时监测 BMC 蛋白质自组装的动态行为方面具有巨大的潜力。
蛋白质-蛋白质相互作用对于 BMC 壳的形成和塑造具有重要意义 [10]。蛋白质浓度也被证明是驱动壳形成的关键因素 [41, 43]。此外,体外溶解度研究表明,溶液中的 pH 值和离子强度会影响 BMCs 的结构稳定性 [17, 27] 以及 BMC 壳蛋白在二维片层形成过程中的组装行为 [37, 41 ]、纳米管 [13, 17] 和纳米笼 [28],让人想起它们对病毒衣壳组装的影响 [44, 45]。我们还发现当 pH> 10 和 <3 或盐浓度 <10 mM 或> 600 mM 时,蛋白质沉淀并且没有形成斑块(未发表的数据)。在这里,我们进一步表明组装趋势和动力学取决于 pH 值和盐浓度。尽管壳蛋白可以在很宽的 pH 范围内自组装,但中性 pH 环境似乎能够增强组装动力学(图 2b)。发现浓度≥ 300 mM的阳离子促进二维片的形成; 400 mM 阳离子似乎是形成大而稳定的单层片的理想选择(图 3)。这些条件与细菌细胞的胞质条件一致,并且在生理上是相关的。例如,在大多数生理相关条件下,E 的 pH 值。大肠杆菌 胞质溶胶约为 7.4–7.8 [46],离子浓度约为 100–400 mM,这对于蛋白质相互作用、蛋白质-配体结合、信号传导、维持膜静电电位和蛋白质跨膜梯度至关重要 [47, 48]。尽管样品与云母底物之间的相互作用如何影响 BMC 蛋白的自组装还有待进一步研究,但 AFM 成像为我们定量分析 BMC 蛋白自组装响应环境变化的动态变化提供了机会。
这里描述的壳片段形成过程中 BMC 蛋白的环境依赖性组装动力学可能代表了它们在整个 BMC 形成中的行为。事实上,3D BMC 结构似乎是自然界设计的动态维持的细胞器。 BMC 具有显着的结构灵活性和异质性;由 AFM 纳米压痕确定的 BMC 壳结构的机械柔软性 [30] 和计算模拟显示的 BMC 组装的非平衡动力学 [49] 突出了 BMC 和稳健病毒组装之间的差异。同样,已经阐明羧基体的生物合成与光和伴侣相关 [50, 51]。最近,已经表明 CcmK3 和 CcmK4 可以形成异六聚体并以 pH 依赖性方式覆盖羧基体壳,这可能提供了一种在高动态微环境中调节羧基体壳渗透性和 CO2 同化的手段 [52]。溶液中的环境条件如何影响 BMC 蛋白的热力学组装的确切机制仍有待研究,例如,结合实验研究和计算模拟。
鉴于 BMC 结构的自组装,人们对工程 BMC 和设计新的基于 BMC 的纳米生物反应器、分子支架和生物技术应用中的生物材料产生了浓厚的兴趣,例如,增强细胞代谢、酶封装、分子递送和治疗.关于 BMC 响应环境变化的结构弹性和可变性的高级知识不仅会为在异源宿主(即 E.大肠杆菌 或植物 [31, 53, 54],但也为调节 2D 纳米材料的形成以及 BMC 壳基蛋白质笼的打开和关闭铺平了道路,从而促进了功能调节和靶向分子递送。以前,我们已经证明了使用遗传修饰方法来操纵壳蛋白界面处的特定接触及其自组装行为的可行性 [12]。这项研究加强了我们在不同环境下评估和操纵 BMC 壳自组装的工具箱。
结论
总之,我们利用 HS-AFM 对不同 pH 和盐条件下 BMC 壳蛋白自组装进行了定量研究。在 400 mM 盐浓度下,更大的单层壳六聚体斑块的形成被促进,中性 pH 导致更高的六聚体自组装动态速率。壳蛋白从六聚体组装到纤维状阵列的组织转变也被可视化。本研究说明环境条件在决定BMC壳蛋白的组织和自组装中起重要作用。
缩写
- BMC:
-
细菌微室
- BMC-H:
-
细菌微室六聚体
- BMC-P:
-
细菌微室五聚体
- BMC-T:
-
细菌微室三聚体
- DSL:
-
动态光散射
- E.大肠杆菌 :
-
大肠杆菌
- EM:
-
电子显微镜
- HS-AFM:
-
高速原子力显微镜
纳米材料