

ROS 和细胞周期停滞在金纳米棒核/银壳纳米结构诱导的基因毒性中的作用
摘要
为了了解银纳米颗粒 (AgNPs) 和银离子在肝脏中诱导的基因毒性,本研究使用了工程金纳米棒核/银壳纳米结构 (Au@Ag NR) 和人源化肝细胞 HepaRG 细胞。氧化应激和细胞周期停滞在 0.4–20 µg mL −1 诱导的 DNA 和染色体损伤中的作用 Au@Ag NR通过彗星试验、γ-H2AX试验和微核试验进行了研究。此外,还分析了 Au@Ag NR 的分布。我们的结果表明,Ag + 和 Au@Ag NR 导致 HepaRG 细胞中的 DNA 裂解和染色体损伤(致畸性),并且保留在细胞核中的 Au@Ag NR 可能进一步释放 Ag + , 加重损害,主要由细胞周期停滞和 ROS 形成引起。结果揭示了细胞内积累与Ag + 之间的相关性 AgNPs 的离子释放和潜在的遗传毒性。
介绍
银纳米粒子 (AgNP) 的大小为 1 至 100 nm,可通过穿透病原体并使其代谢酶的内部巯基失活而表现出广谱的抗菌特性 [1]。它们已在大肠杆菌中证明了有效的抑菌和杀菌作用 , 淋病奈瑟菌 和沙眼衣原体 ,并被广泛用作医用涂料、家用产品 [2] 以及伤口敷料 [3]。令人信服的证据表明,纳米颗粒能够进入细胞核并干扰 DNA 的合成和转录过程 [4]。在我们之前的研究中,我们报道了单次静脉注射 5 mg/kg AgNPs 可以在 Sprague-Dawley 大鼠的骨髓细胞中引入显着的染色体断裂 [5]。单次腹膜内注射 10 毫克/千克或以上的 AgNP 会导致 DNA 和染色体损伤 [6]。花等。 [7] 建议剂量为 50 和 100 μg mL -1 的 AgNPs 可在给药后五分钟内引发 DNA 损伤,突出了快速释放的银 (Ag) 的遗传毒性。考虑到过度暴露的风险,纳米基因毒理学或工程纳米材料的DNA损伤和致癌潜力的研究备受关注[8]。
AgNP 诱导的遗传损伤的主要机制被认为是活性氧化物质的过度产生、炎症和细胞周期紊乱 [9, 10]。正如之前的研究所表明的那样,AgNPs 可以通过氧化损伤直接与 DNA 相互作用 [11] 并在 DNA 水平上干扰间期和染色体水平上的有丝分裂,或者与核蛋白和有丝分裂纺锤体相互作用以干扰细胞周期检查点 [11] 12]。然而,AgNPs 诱导的基因毒性是否部分归因于纳米粒子 [13, 14] 或完全归因于释放的 Ag + 离子仍不清楚[15, 16]。
由于银在组织中不稳定和不间断的释放,因此很难研究 AgNPs 的遗传毒性,从而导致难以定位 AgNPs 并将纳米核与 Ag 区分开来。我们小组最近开发了一种金纳米棒核/银壳(Au@Ag NR)纳米结构,用于研究纳米颗粒引起的毒性[17]。 Au@Ag NR的金核在组织中具有生理先天性,可用作监测Ag + 释放的内标 通过监测 Ag/Au 比率的变化,使用电感耦合等离子体质谱法 (ICP-MS) [18] 测量来自棒的离子。通过这种方法,可以确定毒性的不同来源。以往的研究表明,释放的 Ag + Au@Ag NR壳中的离子导致肾脏氧化损伤,最终导致肾小球的形态变化和过滤功能受损[19]。江等人。 [20] 表明 Au@Ag NR 的颗粒特异性活性和细胞内银离子释放都有助于颗粒细胞的毒性反应。我们还采用 Au@Ag NR 作为模型来研究 AgNPs 的体内遗传毒性潜力,并证明由 Ag 壳和释放的 Ag + 诱导的遗传毒性的主要形式是致畸性,而不是致突变性。上> 离子,而它们的毒性模式没有差异[21]。
肝脏是容易积聚 AgNPs 的主要器官之一,被认为是 AgNPs 诱导的基因毒性的靶器官/组织。我们之前的研究表明,在静脉注射一剂 Au@Ag NR 八周后,SD 大鼠的肝脏中仍有一定量的银 (8.26 ± 3.90 μg/g) 和金 (80.07 ± 64.72 μg/g) [21 ]。在这项研究中,我们尝试使用 Au@Ag NR 在人肝癌衍生的 HepaRG 细胞中确定细胞周期停滞和反应性氧化应激对 AgNP 诱导的染色体和 DNA 损伤的作用。基因毒性试验,包括彗星试验、γ-H2AX 试验和微核试验,与氧化自由基清除剂同时进行,以探测活性氧 (ROS) 在 DNA/染色体损伤中的贡献,而细胞凋亡、细胞周期和相关蛋白决心探索 AgNPs 中断 DNA 合成和复制的机制。此外,通过结合电感耦合等离子体质谱(ICP-MS)和透射电子显微镜(TEM)来区分纳米颗粒和释放的银离子的作用,研究了Au@Ag NR在细胞内的积累和分布。
材料和方法
细胞培养和处理
本研究使用人肝癌细胞系 HepaRG (Thermo Fisher Scientific)。细胞在含有 10% 胎牛血清(FBS,Australia Origin,Gibco)和 1% 青霉素-链霉素-谷氨酰胺溶液(Gibco)的 RPMI 1640 中培养,在 5% CO2 的湿润气氛中,37°C。细胞分别用浓度增加的 Au@Ag NR 处理 24 小时或 72 小时,并根据细胞活力测定估计的 IC50 确定浓度。为了研究 ROS 在遗传毒性中的作用,1 mM N -乙酰-l-半胱氨酸(NAC,Sigma-Aldrich)在用 Au@Ag NR 处理之前应用 1 小时。
ATP 细胞生长/活力测定
将细胞以5 × 10 3 的密度接种于96孔板中 /好。孵育 24 小时后,吸出培养基并将细胞分别暴露于不同浓度的 Au@Ag NR 24 小时或 48 小时。制备了广泛的浓度,并且在一个处理期内每个处理进行了四口井。 Au@Ag NR 的细胞毒性通过三磷酸腺苷 (ATP) 测定法(CellTiter-Glo® 2.0 测定法,Promega)进行检查,该测定法通过定量 ATP 的量来测量细胞代谢活性,ATP 是活细胞中的重要代谢参数。使用 VICTOR Multilabel Plate Reader (2030-0050, PerkinElmer) 检测反映活细胞数量的发光信号,IC50 值估计为 Prism 7 (GraphPad Prism 7,加利福尼亚州,美国)。使用以下等式计算存活率:
$${\text{Viability}}\,{\text{Ratio}}\,\left( \% \right) ={\text{RLU}}_{{{\text{sample}}}} /{ \text{RLU}}_{{{\text{车辆}}}} \times {1}00\%$$其中RLU是相对光单位,表示为四个孔的平均值,RLUvehicle表示未用纳米棒处理的细胞,RLUsample表示用不同浓度Au@Ag NR处理的细胞。
细胞中银和金的浓度测定
使用微波消解系统在硝酸中消解细胞样品。消解后,用含有 1% 硝酸和盐酸的混合物制备样品。溶液中 Ag 和 Au 的量通过 ICP-MS (NexION300X, PerkinElmer) 测定。 TEM 分析用于确定电池中 Au NR 和 Au@Ag NR 的存在。细胞样品在 2.5% 戊二醛和 2% 多聚甲醛的混合物中在 4°C 下固定 2 小时。将细胞沉淀固定并在磷酸盐缓冲液 (pH 7.4) 中冲洗 3 次,然后在 1% 四氧化锇中在 4°C 下固定 2 小时。随后将样品在蒸馏水中漂洗 3 次,并在不同浓度的乙醇(分别为 50%、70%、90% 和 100% 乙醇)中一个接一个地脱水 15 分钟。随后,将 1:1 和 1:3 稀释的环氧丙烷在 20-26°C 下施加到树脂上 2 小时。聚合通过在 35°C 16 小时、45°C 8 小时、55°C 14 小时和 65°C 48 小时分级加热来进行。超薄切片用醋酸双氧铀和柠檬酸铅染色 25 分钟,并通过透射电子显微镜(H-7650,HITACHI,日本)进行分析。
常规和改良彗星试验
将细胞以2 × 10 5 的密度接种于12孔板中 /well 或 3 × 10 5 /well 分别进行 24 小时或 72 小时处理。将浓度为 200 μmol 的过氧化氢 (H2O2) 作为阳性对照暴露于细胞中一小时。对于每个样品,为常规处理和甲酰氨基嘧啶糖基化酶 (Fpg) 处理制备两个孔。如前所述[21],常规彗星试验是在碱性条件下(pH> 13)进行的。对于 Fpg 处理的孔,在 DNA 展开程序之前应用额外的 Fpg 处理,并将载玻片浸入酶缓冲液(0.1 M KCl、0.5 mM EDTA、40 mM HEPES、0.2 mg.mL -1 BSA) 3 次,每次 5 分钟。 Fpg (New England Biolabs, Inc., UK) 用酶缓冲液以 1:50,000 稀释。将 100 毫升稀释酶的等分试样加入显微镜载玻片上的每个凝胶中,并在 37°C 的湿度室中孵育 30 分钟。其余步骤同常规处理。彗星试验一式三份进行。使用 Nikon Eclipse 80i 荧光显微镜(Nikon,Tokyo,Japan)对每个样本至少 50 个细胞进行独立评分,同时使用 Komet 6.0(Andor Technology,Belfast,UK)分析尾部和橄榄尾中 DNA 百分比的中值每个样品的矩(OTM)。
通过流式细胞术和高内涵筛选鉴定 γ-H2AX 病灶
为了使用流式细胞术进行定量,将细胞以 2 × 10 5 的密度接种在 12 孔板中 /well 或 3 × 10 5 / 孔分别进行 24 小时或 72 小时处理,而对于高内涵筛选试验,将细胞以 6 × 10 3 的密度接种在 96 孔板中 /well 或 1 × 10 4 /well 分别进行 24 小时或 72 小时处理。作为阳性对照,2 μM 甲磺酸甲酯(MMS,Sigma-Aldrich)与细胞平行应用一小时。在 Tris 缓冲盐水 (TBS) 中冲洗细胞并在室温下用 4% 多聚甲醛固定 15 分钟。用 TBS 洗涤后,将细胞与 50 μL 冰冷的甲醇在 - 20°C 下孵育 30 分钟。细胞在 TBS 中进一步冲洗 3 次,并应用封闭剂(含有 0.3% Triton X-100 和 10% 山羊血清的 TBS)1 小时。用封闭剂将一抗(小鼠抗磷酸化 H2AX Ser139,Millipore)稀释至 1:200,并与细胞在 4°C 下孵育过夜。然后再次用 TBS 冲洗板 3 次,随后加入用封闭剂以 1:20 比例稀释的二抗(Alexa Fluor 488 山羊抗小鼠,Life Technologies)。样品在室温下避光 1 小时,2 μg mL -1 将 (20 μL/孔) DAPI (Invitrogen) 添加到每个孔中。使用流式细胞术(FACSCalibur,BD Bioscience,NJ,USA)或高内涵分析系统(Operetta CLS,PerkinElmer)测量荧光。对于流式细胞术测定,分析了每组至少 10,000 个细胞的数据,并且实验一式三份进行;高内涵分析每孔20个视野,每组至少5个孔。
细胞分裂阻滞微核细胞组 (CBMN-cyt) 检测
CBMN-cyt 根据 Fenech 等人描述的程序进行。 [22]。将细胞以2 × 10 5 的密度接种于12孔板中 /well 或 3 × 10 5 /well 分别进行 24 小时或 72 小时处理。 0.2 μg mL −1 丝裂霉素 C(MMC,Tokyo Chemical Industry Co, Ltd. Japan)暴露于作为阳性对照的细胞 24 小时。 3 μg mL −1 在处理 24 或 72 小时后应用细胞松弛素 B 以阻断胞质分裂过程,并在 40 小时后收获细胞。样品用预热 0.075 mol L −1 低渗后用 5% Giemsa 染色 KCl 并用 3:1 的甲醇和乙酸混合物固定。每组设3个平行孔,每孔至少检测1000个双核细胞。
MDA、总GSH和SOD含量测定
细胞在12孔板中培养,密度为5 × 10 5 /well 或 3 × 10 5 /well 分别进行 24 小时或 72 小时处理。随后,收获细胞并用磷酸盐缓冲盐水 (PBS) 冲洗 3 次。使用基于硫代巴比妥酸的方法(南京建成生物工程研究所,南京,中国)测定细胞匀浆中丙二醛(MDA)的量。分别使用总谷胱甘肽定量和 SOD 测定试剂盒(Dojindo Molecular Technologies, Inc. Kumamoto, Japan)测定总谷胱甘肽 (GSH) 和超氧化物歧化酶 (SOD) 的量。使用VICTOR Multilabel Plate Reader(2030-0050,PerkinElmer)测量每孔的光密度(O.D)。
细胞周期流式细胞术分析
细胞在6孔板中培养,密度为1 × 10 6 /well 或 5 × 10 5 / 孔分别进行 24 小时或 72 小时处理,随后在 4°C 下用 70% 乙醇固定过夜。样品用 PBS 冲洗 3 次,并用 PI/Rnase 染色缓冲液(BD Biosciences)在室温下染色 15 分钟。通过使用具有 FL2 面积与 FL2 宽度的区域来确定 20,000 个细胞中处于 G0/G1、S 和 G2/M 期的细胞群。采用流式细胞术(FACSCalibur,BD Bioscience,NJ,USA)和FlowJo(BD Bioscience)进行分析,实验一式三份。
细胞凋亡的流式细胞分析
细胞在6孔板中培养,密度为1 × 10 6 /well 或 5 × 10 5 /well 分别进行 24 小时或 72 小时处理。随后用 PBS 冲洗两次并用 500 μL 1 × 结合缓冲液(FITC 膜联蛋白 V 细胞凋亡检测试剂盒 I,BD Bioscience)稀释,以将悬浮液调整至约 1 × 10 6 细胞/mL,随后将 100 μL 稀释液与 5 μL FITC Annexin V 和 5 μL PI 混合。样品在室温下染色 15 分钟,并使用流式细胞术(FACSCalibur,BD Bioscience,NJ,USA)和 FlowJo 分析至少 10,000 个细胞以确定早期和晚期凋亡的细胞群(BD 生物科学)。实验一式三份进行。
蛋白质印迹分析
细胞在 75 cm 2 密度为 1 × 10 7 的烧瓶 /well 和 6 × 10 6 /well 分别进行 24 小时和 72 小时的处理。用含有蛋白酶抑制剂(PMSF)的RIPA裂解缓冲液裂解细胞,并使用BCA蛋白质定量试剂盒(Beyotime Biotechnology,China)测定蛋白质浓度。在通过在 95°C 下加热 3 分钟进行变性之前,使用 RIPA 裂解缓冲液调整样品的浓度。通过在 12% SDS 聚丙烯酰胺凝胶上电泳分离蛋白质样品并转移到硝酸纤维素膜 (Millipore) 上。膜用 5% 脱脂牛奶封闭 30 分钟,并与初级 p53(SC-137174,Santa Cruz)、p21(SC-6246,Santa Cruz)和 β-肌动蛋白(sc-47778,Santa Cruz)和二抗一起孵育山羊抗小鼠 IgG(H+L)-HRP(SE131, solabio),分别。使用增强化学发光(ECL)方法观察样品中靶蛋白的表达水平,并通过ImageJ系统(National Institutes of Health)进行分析。
统计分析
数据表示为平均值 ± SEM。用单向方差分析(ANOVA)检验阴性对照组和治疗组之间差异的统计显着性,然后用SPSS(第22版,IBM,Armonk,NY,USA)进行Dunnett多重比较检验,并考虑数据P 的统计显着性 <0.05。图形使用 Windows 版 GraphPad Prism 7(GraphPad Software, La Jolla, CA, USA)制作。
结果
Au NR 和 Au@Ag NR 的表征
金纳米棒(Au NRs)、金纳米棒核和银壳纳米结构(Au@Ag NR)的设计、制备和表征如前所述[21]。简而言之,Au NRs 的平均直径和长度为 15.0 ± 2.5 nm,66.7 ± 2.5 nm,Au@Ag NRs 的平均直径和长度为 26.2 ± 3.0 nm,72.7 ± 8.9 nm。 Ag 壳厚度约为 5 纳米。分散在水中的 PDDAC 包覆的 Au NRs 和 Au@Ag NRs 的 zeta 电位分别为 37.7 ± 1.6 mV 和 52.5 ± 1.4 mV。制备的 Au@Ag NR 的 Ag/Au 重量比估计为 2.3。表征结果如图1所示。
<图片>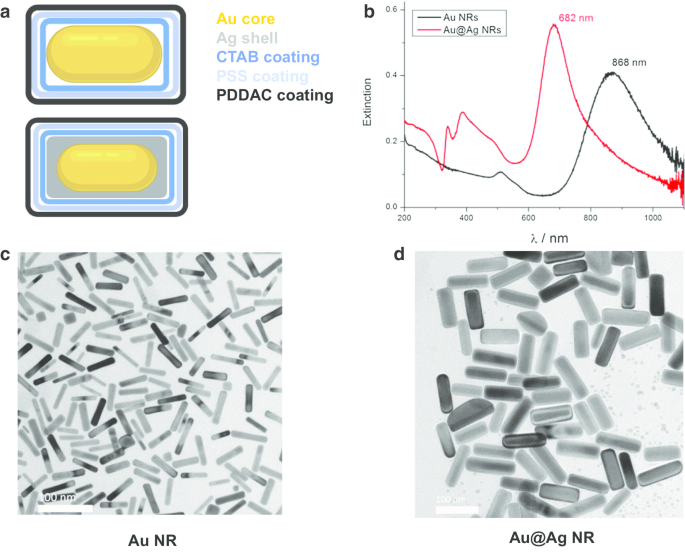
Au NR 和 Au@Ag NR 的表征。 一 Au NR和Au@Ag NR的结构图; b 分散在水中的Au NR和Au@Ag NR的UV-Vis-NIR消光光谱; c Au NR的代表性TEM图像; d Au@Ag NR的代表性TEM图像
细胞活力
Au@Ag NR 对 HepaRG 细胞的细胞毒性通过 ATP 活力测定(表 1)进行研究,细胞暴露于 Au@Ag NR 中 24 或 48 小时,浓度从 0.125 到 160 μg mL -1 .暴露 24 小时和 48 小时后,Au@Ag NR 以时间和剂量依赖性方式诱导显着的细胞毒性作用,在 20 µg mL -1 时的存活率 IC50 百分比 和 6 µg mL −1 , 分别由软件 GraphPad Prism 7.0 拟合。考虑到整体细胞毒性,将处理时间调整为 24 小时和 72 小时,而应用的浓度确定为 0.8 µg mL -1 , 4 µg mL −1 和 20 µg mL −1 .此外,加入了 Au NR 作为惰性对照,AuNR 组中的 Au 含量与 20 µg mL -1 相同 Au@Ag NR,为 16 µg mL −1 .相比之下,Au@Ag NR + NAC 组采用 1 mM NAC 预处理作为氧化应激反应的对照(Au@Ag NR 的浓度为 20 μg mL -1 ).
Au NR 和 Au@Ag NR 的细胞分布
HepaRG 细胞中 Au 和 Ag 含量的分布通过 ICP-MS 进行分析。如表 2 和表 3 所示,Ag 含量呈剂量依赖性增加。然而,抗氧化剂N -乙酰-l-半胱氨酸 (NAC) 作为自由基清除剂可能会限制纳米颗粒的细胞摄取,因为即使 Au@Ag NR 浓度相同(20 µg mL -1 ) 应用于该组。 Ag/Au 比率从 24 小时下降到 72 小时表明 Ag + 的持续释放 来自 Au@Ag NR 的壳。此外,Ag 的细胞摄取量远高于 Au(表 4)。此外,TEM 数据显示,大部分 Au NR 和 Au@Ag NR 作为团聚体保留在细胞中。在Au NR或Au@Ag NR暴露的细胞内,纳米棒结构清晰可见,但未进入细胞核(图2)。
<图片>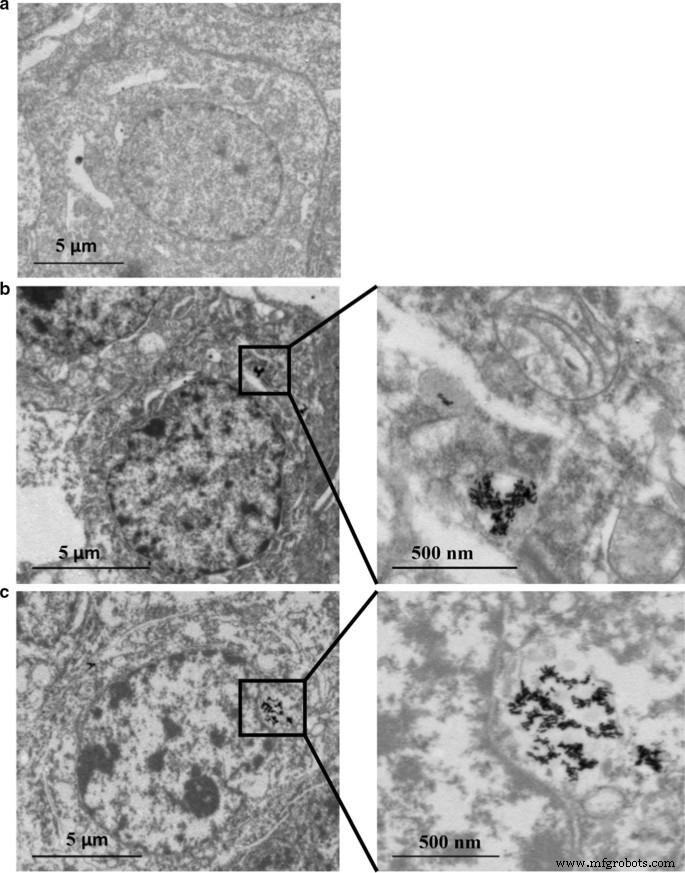
Au NR 和 Au@Ag NR 内化:在 80 kV 下通过 TEM 在 16 μg mL −1 中暴露 24 小时后的 HepaRG Au NR 和 20 μg mL −1 Au@Ag NR。 一 车辆控制; b 金NR; c Au@Ag NR
DNA 损伤
Au@Ag NR 引发的 DNA 损伤通过彗星试验和 γH2AX 试验进行评估(图 3)。从彗星试验中观察到,0.8 至 20 µg mL -1 Au@Ag NR 可能会导致显着的 DNA 损伤。暴露于 Au@Ag NR 24 或 72 小时后,细胞的尾 DNA 百分比和 OTM 均以时间和浓度依赖性方式增加。此外,在用 20 µg mL -1 处理的细胞中观察到与氧化应激诱导相关的 DNA 损伤 Au@Ag NR 通过 Fgp 酶修饰彗星试验(图 3a、b)。为了评估与癌症发生具有更高相关性的双链断裂程度,分析了 γ-H2AX 阳性细胞和 γ-H2AX 阳性细胞中的平均荧光强度。暴露于 Au@Ag NR 24 小时后,γ-H2AX 阳性细胞的组间没有差异。然而,4 μg mL −1 Au@Ag NR 组在处理 72 小时后引起显着增加。与载体对照相比,72 小时后所有 Au@Ag NR 组的荧光强度均显着增加(图 3c-e,P <0.05).
<图片>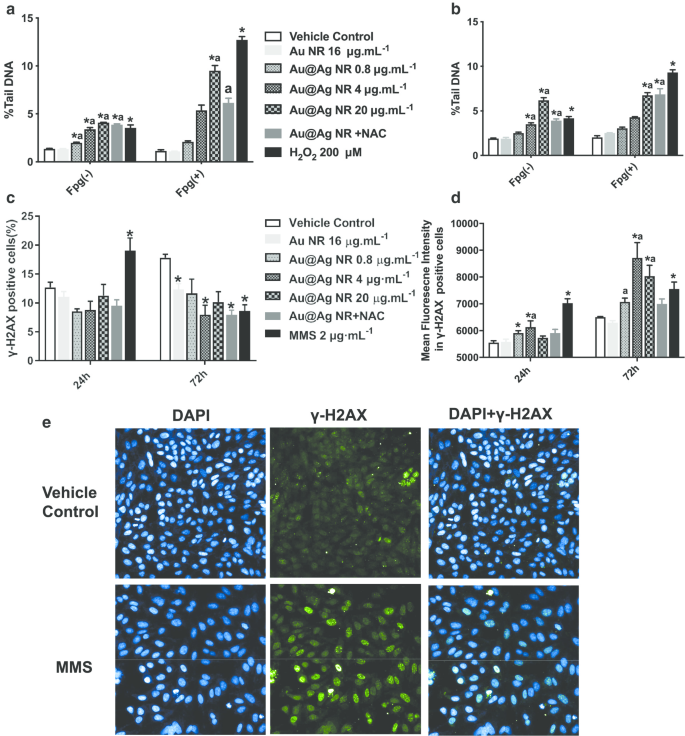
Au@Ag NR 诱导的 DNA 损伤。 HepaGR 细胞暴露于不同浓度的 Au@Ag NR(0.8 至 20 μg mL -1 ) 分别为 24 小时和 72 小时。 一 暴露于 Au@Ag NR 24 小时后的尾 DNA 平均百分比; b 暴露于 Au@Ag NR 72 小时后的平均尾 DNA 百分比; c 使用流式细胞术估计具有 γ-H2AX 病灶的阳性细胞的百分比; d 使用免疫荧光染色估计具有 γ-H2AX 病灶的细胞的平均荧光强度。 * P <0.05 与车辆对照; 一 P <0.05 与 Au NR。 2 μM mL −1 MMS作为阳性对照
染色体损伤
微核的形成是识别染色体损伤的重要生物标志物,染色体损伤是比 DNA 断裂对遗传物质更严重的损伤。如图 4c 所示,对含有微核的双核细胞的比例进行评分。 Au@Ag NR 以浓度依赖的模式增加了微核的形成。暴露 24 小时后,在用 4 μg mL -1 处理的细胞中观察到的微核比率 Au@Ag NR 和 20 μg mL −1 Au@Ag NR分别为1.133 ± 0.145%和1.567 ± 0.318%,均显着高于载体对照组。暴露 72 小时后,用 4 μg mL -1 处理的细胞中微核的比例 Au@Ag NR为1.767 ± 0.233%,明显高于赋形剂对照组;用 20 μg mL −1 处理的细胞中微核的比例 Au@Ag NR 为 2.167 ± 0.252%,显着高于载体对照组和 16 μg mL -1 Au NR 组(0.700 ± 0.153%)。相比之下,用 20 μg mL -1 处理的细胞之间没有发现差异 Au@Ag NR + NAC和载体对照,表明ROS参与了Au@Ag NR诱导的染色体断裂。
<图片>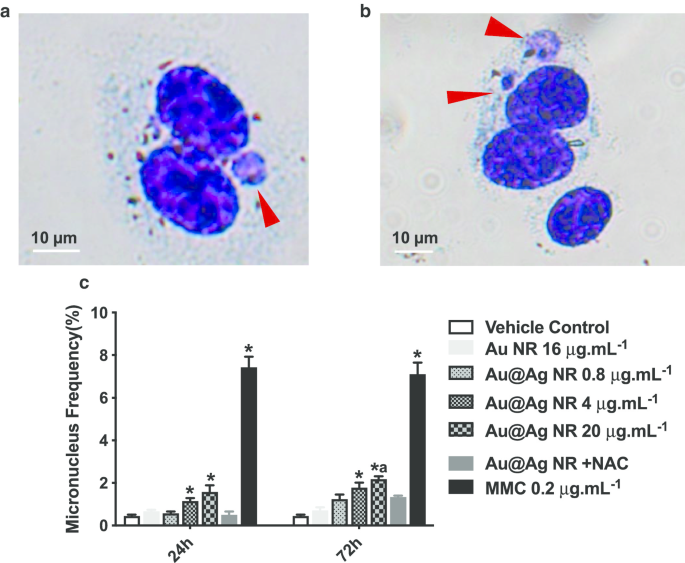
Au@Ag NR 诱导的染色体损伤。 HepaGR 细胞暴露于不同浓度的 Au@Ag NR -1 至 20 μg mL −1 24 小时和 72 小时。 一 , b 微核的代表性图像(红色箭头); c 微核频率 (%)。 * P <0.05 与车辆对照; 一 P <0.05 与 Au NR。 0.2 μg mL −1 丝裂霉素C作为阳性对照
Au@Ag NR 对 ROS 形成的影响
为了进一步探索 ROS 形成在 Au@Ag NR 诱导的 DNA 和染色体损伤中的作用,估计了 MDA、GSH 和 SOD 水平。 MDA 形成显着增加 (P <0.05) 在暴露于 20 μg mL -1 后观察到 24 小时和 72 小时的 Au@Ag NR(图 5a)。此外,暴露于 Au@Ag NR 的细胞中 GSH 和 SOD 水平显着降低(P <0.05) 以时间和浓度依赖的方式。这些结果表明,Au@Ag NR 的暴露产生了氧化和抗氧化之间的不平衡(图 5b、c)。
<图片>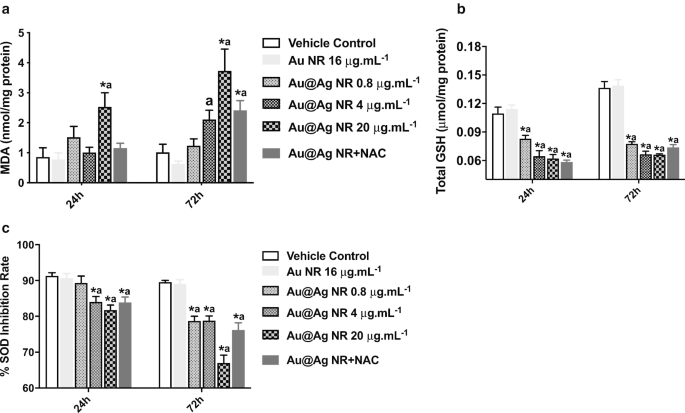
Au@Ag NR对ROS形成的影响。 HepaGR 细胞暴露于不同浓度的 Au@Ag NR -1 至 20 μg mL −1 24 小时和 72 小时。 一 MDA 级别; b GSH水平; c SOD 水平。 * P <0.05 与车辆对照; 一 P <0.05 vs Au NR
Au@Ag NR 对细胞周期和细胞凋亡的影响
暴露于 Au@Ag NR 72 小时后,在 4 μg mL -1 中观察到 G2/M 期细胞数量增加 Au@Ag NR,20 μg mL −1 Au@Ag NR和Au@Ag NR + NAC组的比例分别为32.63% ± 1.77%、32.267% ± 2.17%和32.967% ± 4.25%(图6a,b)载体对照组(22.37% ± 0.92%)。同时,Au@Ag NR 暴露 72 小时后可观察到细胞凋亡,20 μg mL -1 处理细胞的晚期凋亡率 Au@Ag NR 和 20 μg mL −1 Au@Ag NR + NAC 分别为 78.90 ± 1.19% 和 70.20 ± 4.50%(图 6c、d)。 Au@Ag NR诱导的晚期细胞凋亡多于早期细胞凋亡,NAC处理可降低Au@Ag NR引发的晚期细胞凋亡率。
<图片>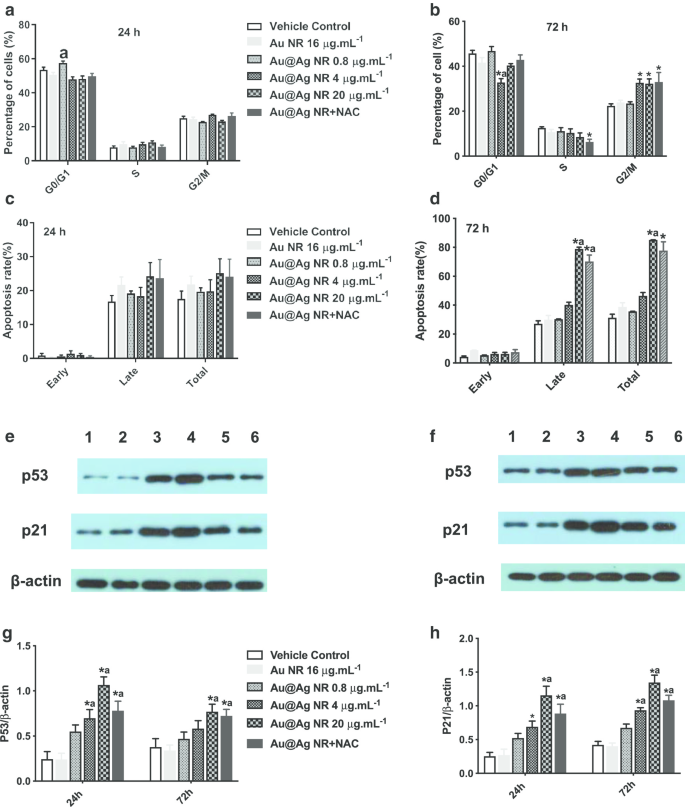
Au@Ag NR对细胞周期和细胞凋亡的影响。 Au@Ag NR对细胞周期的影响(a , b ) 和细胞凋亡 (c , d ) 分别暴露 24 小时和 72 小时后;不同组HepaRG细胞中p53和p21表达水平的代表性数据(e , f 车道1:车辆控制;泳道 2:Au NR;泳道3:Au@Ag NR + NAC;泳道 4:Au@Ag NR 20 μg mL −1 ;泳道 5:Au@Ag NR 4 μg mL −1 ;泳道 6:Au@Ag NR 0.8 μg mL −1 );不同组中 p53 和 p21 与 β-actin 的平均相对表达水平总结在 (g , f ). * P <0.05 与车辆对照; 一 P <0.05 vs Au NR
通过蛋白质印迹检测 p21 和 p53 的表达水平,并观察到类似的模式。用 4 μg mL -1 处理的细胞中 p53 和 p21 的表达水平 和 20 μg mL −1 Au@Ag NR 显着增加 (P <0.05) 并且在用 20 μg mL -1 处理的细胞中显着降低 Au@Ag NR 和 NAC (P <0.05,与 20 μg mL 相比 -1 Au@Ag NR 组,图 6e-h)。已知p53蛋白是响应DNA损伤而介导G2/M检查点激活的核心分子,并且p21被认为是p53依赖性细胞周期抑制剂。因此,Au@Ag NR可以干扰DNA复制并通过细胞周期停滞阻碍DNA修复。
讨论
目前发布的Ag + 的作用 和 AgNPs 在产生遗传毒性方面的作用尚不清楚。我们小组 [21] 和其他人 [13] 之前的研究表明,虽然 Ag + 是引入毒性的主要来源,但纳米颗粒也可能是剧毒的。例如,AgNPs 可以通过诱导羟基自由基的形成而导致基因毒性 [13]。 Further, more severe chromosome damage, oxidative stress and apoptosis were introduced by AgNP compared to Ag + alone [23], suggesting that different pathways might be involved. We employed Au@Ag NR as a model material to understand the forms and distributions of AgNPs in cells, and the amounts of intracellular Ag and Au were determined by ICP-MS. The Ag/Au weight ratio of prepared Au@Ag NR was estimated as 2.3. However, after a 24-h exposure, it sharply increased to 16.5 in the cells treated with Au@Ag NR, suggesting that large amount of Ag was released from the shell of Au@Ag NR within that period. When the exposure period of Au@Ag NR was extended to 72 h, the Au/Ag weight ratio was decreased to 1.7, indicating that the Ag + was released from the cell and the nanorod was the major form of Au@Ag NR in the cell at that stage. Therefore, it could be deduced that once the Au@Ag NR entered the cell, Ag + rapidly dissolved from its shell within 24 h and gradually released to the extracellular environment, while the Au@Ag NR itself retained in the cell for a longer period.
Oxidative stress is deemed as one of the most important toxicological mechanisms of nanoparticles [24]. N-acetylcysteine (NAC) is a thiol, a mucolytic agent and precursor of l-cysteine which reduced glutathione. NAC is also a source of sulfhydryl groups in cells and exerts the ROS scavenger activity by interacting with OH · and H2O2[25]. In this study, the GSH and SOD levels were significantly decreased after exposure to Au@Ag NR, while the MDA level increased in a concentration- and time-dependent manner, indicating that the Au@Ag NR introduced the oxidative stress in the cells.
The potentials of Ag and Au@Ag NR in interfering with the genetic materials were further investigated by a series of genotoxicity assays. It is noteworthy that co-culturing the NAC with Au@Ag NR could ameliorate the ROS formation, which in turn supports the participation of oxidative stress in the genotoxicity triggered by Au@Ag NR. In this study, comet and γ-H2AX assays were performed to confirm that Au@Ag NR could interact with DNA and induce certain DNA damage, and the repair endonuclease Fpg was included in the comet assay to identify the oxidative DNA damage [26]. The Fgp could recognize oxidized pyrimidines and remove oxidized purines, e.g., 8-hydroguanine, so as to create apurinic or apyrimidinic sites that could introduce gaps in the DNA strands. The oxidative stress-induced DNA breakage could be determined subsequently by another comet assay [27]. The further DNA breakage detected by the additional Fgp in the comet assay suggested that the Au@Ag NR could cause DNA damage. Mei et al. [28] observed that 5-nm-sized AgNPs induced oxidative lesion-specific DNA damage by employing the hOGG1, EndoIII and Fpg endonucleases in the comet assay.李等人。 [29] also suggested that both PVP- and silica-coated AgNPs (15–100 nm and 10–80 nm, respectively) could lead to a significant increase in DNA breakage in mice hepatocytes in the presence of hOGG1and EndoIII. The formation of γ-H2AX foci, which represents an early cellular response to genotoxic stress, is the most sensitive and specific biomarker for detecting DSBs [30]. As demonstrated in this study, γ-H2AX foci in cells exposed to Au@Ag NR were markedly increased after 24 h, and a further increase could be observed after 72 h. The reduction in the 20 µg mL −1 group might be due to the cytotoxicity to the HepaRG cells at higher concentration. Similar results were observed for AgNPs with different coatings [31, 32]. Further, our results suggest that Au@Ag NR could induce chromosome damage in HepaRG cells, as the micronucleus rates were significantly increased. This is consistent with previous studies, where AgNPs-induced increased micronucleus rate was reported in HaCaT and TK6 cells [33]. In contrast, the addition of oxidative radical scavenger NAC could inhibit the formation of micronucleus induced by Au@Ag NR. Taken together, these data suggest the participation of oxidative stress in AgNP-introduced clastogenicity risk in vitro.
Previous studies have investigated the cell cycle arrest and cytotoxicity induced by AgNPs [33,34,35]. With prolonging the exposure time, the impact of AgNPs on cell cycle and apoptosis might be enhanced and in turn aggravate the cytotoxicity and genotoxicity. Usually, the cell cycle checkpoints (e.g., G2/M) were initiated by cells when experiencing DNA damage, and this mechanism serves to prevent the cell from entering mitosis (M phase). The G2/M cell cycle arrest indicates that an increasing percentage of cells is hindered in G2 phase for DNA repairing. Cells experiencing successful DNA repairing would further proceed to mitosis; however, for those with fatal damages, irreversible G2/M cell cycle arrest and cells apoptosis would take place [36]. We observed that Au@Ag NR could arrest the majority of HepaRG cells in G2/M phase, induce late cell apoptosis and increase the expression levels of p53 and p21, which are important proteins associated with the regulation of cell cycles [37]. As p53 could also induce apoptosis, when the DNA cannot be repaired properly [38], the p21 might indirectly participate in cell apoptosis by cell cycle arrest in a p53-dependent pathway via down-regulating the nuclear protein ICBP90 for DNA replication and cell cycle regulation [39]. Furthermore, apoptosis and a G2/M arrest induced by activation of the p53/p21 system have been reported in HepG2 cells following the administration of garlic extracts [40]. Thus, it could be inferred that the oxidative stress-triggered DNA/chromosome damages might facilitate the expression of p53 and p21, which subsequently induces cell cycle arrest. Extending the exposure period of Au@Ag NRs to the DNA/chromosome during replication may further aggravate the genotoxicity or apoptosis.
Conclusion
Genotoxicity induced by AgNPs may be attributed to the oxidative stress induced by the nanoparticles as well as the released ions [41]. This study employed Au@Ag NR as a model to determine the distribution and release behavior of Ag after the nanoparticles enter into the cells. Considering the disparate forms of Au@Ag NR in the cell, after its exposure the Ag + was rapidly dissolved from the silver shell. Ag + and Au@Ag NR could introduce cytotoxicity and genotoxicity (clastogenicity) in the cells, and the Au@Ag NR retained in the nucleus may further release Ag + to aggravate the damage, which are mainly caused by cell cycle arrest and ROS formation (summarized in Fig. 7). Collectively, these data reveal the correlation between the intracellular accumulation, Ag + release as well as the potential genotoxicity of AgNPs.

Schematic diagram of the possible mechanism of genotoxicity introduced by AgNP in vitro
数据和材料的可用性
All data and materials are available without restriction.
纳米材料
- 钨银合金与钨铜合金的区别
- 负载 ICA 的 mPEG-ICA 纳米颗粒的制备及其在治疗 LPS 诱导的 H9c2 细胞损伤中的应用
- 研究乳液液滴尺寸和表面活性剂在基于界面不稳定性的胶束纳米晶体制造过程中的作用
- TiO2 中金纳米粒子分布对染料敏化太阳能电池光学和电学特性的影响
- Au@TiO2 蛋黄-壳纳米结构的制备及其在亚甲基蓝降解和检测中的应用
- 用 6-巯基嘌呤和神经元穿透肽修饰的金纳米颗粒促进 SH-SY5Y 细胞生长
- CoFe2O4/Fe3O4 和 Fe3O4/CoFe2O4 核/壳纳米颗粒中的深刻界面效应
- 桔梗皂苷(桔梗)用于金和银纳米颗粒的绿色合成
- 用于新型隐球菌诊断和光热治疗的抗体偶联二氧化硅修饰金纳米棒:体外实验
- 评估由食用菌根真菌 Tricholoma crassum 合成的蛋白质包覆金纳米粒子的抗菌、凋亡和癌细胞基因传递特性
- 无镉 Cu 掺杂 ZnInS/ZnS 核/壳纳米晶体:可控合成和光物理特性
- 从石榴皮提取物中合成的银纳米粒子的抗菌和细胞毒性作用