

Nec-1 通过 RIP1 减弱二氧化钛纳米材料对 Sh-Sy5y 细胞的神经毒性
摘要
二氧化钛纳米材料由于其优异的理化特性而被应用于众多领域,进而对人类健康构成潜在威胁。最近,大量的体内研究表明,二氧化钛纳米粒子 (TNPs) 在暴露后可以通过各种途径转运到动物大脑中。吸收的 TNP 会在大脑中积累,并可能干扰神经元细胞,导致大脑功能障碍。体外研究证实了 TNP 的神经毒性。 TNPs 的神经毒性机制尚不清楚。坏死性凋亡是否与 TNP 的神经毒性有关尚不清楚。因此,我们进行了一项体外研究,发现 TNPs 以剂量依赖性方式诱导 SH-SY5Y 细胞的炎症损伤,而 necrostatin-1 (Nec-1) 预处理可以减轻这种损伤。由于据报道受体相互作用蛋白激酶 1 (RIP1) 是 Nec-1 的靶标,我们通过 siRNA 使其沉默。我们将突变型和野生型细胞暴露于 TNP 并评估炎症损伤。沉默 RIP1 表达可抑制 TNPs 暴露诱导的炎症损伤。总之,Nec-1 通过 RIP1 改善 TNP 的神经毒性。然而,还需要更多的研究来综合评估TNPs的神经毒性与RIP1之间的相关性。
介绍
由于其优异的物理化学性质,二氧化钛纳米材料被合成 [1] 并广泛用于各种用途,如化妆品 [2]、工业领域 [3, 4] 和医疗领域 [5]。然而,这种广泛使用可能对人类健康构成巨大威胁[6]。一旦暴露,大部分二氧化钛纳米粒子 (TNP) 通过吸入和摄入进入人体 [6]。呼吸系统 [7]、消化系统 [8] 和心血管系统 [9] 都可能被吸收的 TNP 中断。同样,大脑作为中枢神经系统最重要的部分,也会受到干扰,因为循环中的TNPs可以穿透血脑屏障,吸入的NPs可以通过嗅觉途径进入大脑[10] .一旦 TNPs 进入大脑,它们可能会在那里积聚并对大脑造成损害,从而导致功能障碍。大脑的损伤通常是不可逆的和严重的,因此应调查任何潜在的损伤原因[11]。
虽然没有流行病学研究探讨 TNPs 暴露与脑部疾病之间的关联,但大量的体内和体外研究证实了 TNPs 的神经毒性 [12]。此外,主要的重点是研究以揭示潜在的机制。为此,我们之前回顾了目前已知的 TNPs 神经毒性分子机制,发现程序性细胞死亡 (PCD) 过程,如细胞凋亡和自噬,与 TNPs 的神经毒性有关 [13]。
坏死性凋亡,也称为调节性坏死,是另一种类型的 PCD。与依赖半胱天冬酶的细胞凋亡不同,坏死性凋亡是一种受体相互作用蛋白激酶 1/3 (RIP1/RIP3) 依赖的。激活的 RIP1 招募 RIP3 形成坏死体,从而激活混合谱系激酶结构域样蛋白 (MLKL) 以启动坏死性凋亡 [14]。坏死性凋亡可以调节细胞死亡和神经炎症 [15, 16],这两者都与 TNP 的神经毒性有关。因此,我们假设坏死性凋亡与TNPs引起的神经毒性有关。
为了验证我们的假设,我们进行了一项体外研究,以探讨坏死性凋亡在 TNP 的神经毒性中的作用。在这项研究中,在 TNPs 暴露后测量了细胞活力、LDH 渗漏和炎症细胞因子(TNF-α、IL-1β、IL-6 和 IL-8)。首先,用不同浓度的 TNP 培养 SH-SY5Y 细胞。其次,将细胞暴露于有或没有 Nec-1 的 TNP,Nec-1 是一种有效的坏死性凋亡抑制剂。第三,可以被 Nec-1 抑制的 RIP1 基因表达使用 siRNA 沉默,然后用 TNP 处理突变和野生型细胞。本研究为全面了解TNPs神经毒性的分子机制提供了依据。
结果
TNP 抑制细胞活力
为了验证 TNPs 对 SH-SY5Y 细胞的细胞毒性,我们首先使用 CCK8 测定评估了细胞活力。将细胞以 5 至 160 μg/mL 的 TNP 浓度培养 24、48 和 72 小时。如图 1 所示,暴露 24 小时后,用 5、10、20 和 40 μg/mL 处理的细胞的细胞活力保持不变。当细胞处理 48 和 72 小时后,仅 5 μg/mL 组的细胞活力保持不变(p =0.4507 在 48 小时和 p =0.1002 在 72 小时)。此外,在 48 天后,用 80 μg/mL TNPs 处理的细胞的细胞活力显着降低 (p =0.0007) 和 72 小时 (p =0.0008)。基于这些结果,我们得出结论,TNPs 以剂量和时间依赖性方式降低细胞活力(数据未显示),表明长期接触毒性更大。
<图片>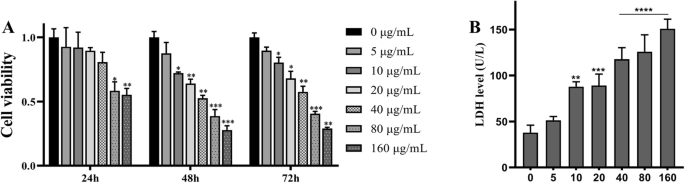
不同浓度的 TiO2-NPs 暴露对 SH-SY5Y 细胞在 24、48 和 72 小时的细胞活力和 72 小时的 LDH 泄漏的影响。 (与对照组相比, * p <0.05, ** p <0.01, *** p <0.001, **** p <0.0001)
TNPs 破坏膜完整性
Nest,我们分析了 TNP 在暴露 72 小时后对膜完整性的毒性影响。通过测量 LDH 渗漏来评估膜完整性。如图 1b 所示,用高于 5 μg/mL 的剂量 TNP 处理的细胞中 LDH 水平显着增加。在用 40、80 和 160 μg/mL 处理的细胞中观察到 LDH 产量增加(p <0.0001)。这些结果表明TNPs以剂量依赖性方式增加LDH水平,这与CCK8检测相似。
TNP 暴露促进炎症
使用 ELISA 分析 TNPs 暴露后的炎症反应。细胞用不同浓度的 TNPs 处理 72 小时后,测量 TNF-α、IL-1β、IL-6 和 IL-8 的水平。如图 2 所示,所有 TNPs 处理的细胞中 IL-8 的分泌均上调;在用高于 5 μg/mL 的剂量处理的细胞中,TNF-α、IL-1β、IL-6 和 IL-8 的水平均匀升高(p <0.01)。此外,在用高于 10 μg/mL 的剂量处理的细胞中炎症显着更高(p <0.0001)。总之,TNPs以剂量依赖性方式增强炎症。
<图片>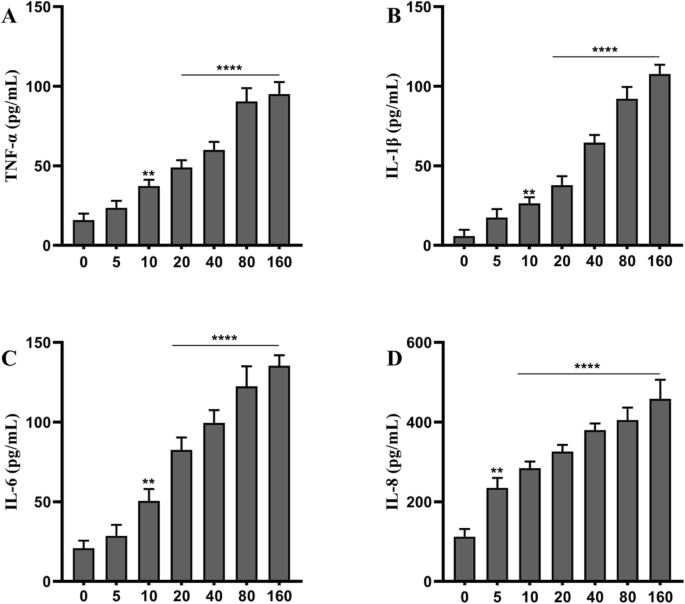
不同浓度的 TiO2-NPs (μg/mL) 在 72 小时时对 SH-SY5Y 细胞炎症的影响。 (与对照组相比, **** p <0.0001)
我们的结果表明,TNPs 以剂量依赖性方式诱导炎症损伤。我们在接下来的实验中采用80 μg/mL的TNPs浓度和72 h的暴露时间来探讨坏死性凋亡在TNPs神经毒性中的作用。
Nec-1 协同治疗抑制 TNP 的神经毒性
为了分析坏死性凋亡在炎症损伤中的作用,我们用 Nec-1(一种坏死性凋亡抑制剂)和 TNP 共同处理细胞。如图 3a 所示,SH-SY5Y 细胞暴露于 TNPs 或 TNPs+Nec-1(1、5、10、15 或 20 μM),我们发现 Nec-1 显着改善了 TNPs 诱导的减少细胞活力 (10, 15, 20 μM) (p <0.0001)。由于 15 μM 和 20 μM 处理组的细胞活力不高于 10 μM 处理组(p =0.6643 和 p =0.6292),我们用 10 μM Nec-1 共同处理细胞以分析对膜完整性的影响。
<图片>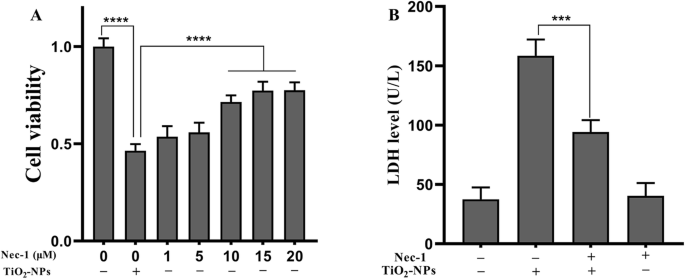
Nec-1 对 TNPs 暴露后细胞活力和 LDH 的影响。 ( *** p <0.001, **** p <0.0001)
用TNPs或TNPs+Nec-1培养细胞后,我们发现TNPs+Nec-1组的LDH水平远低于单用TNPs组(p =0.0005)。此外,单独使用 Nec-1 处理不会增加 LDH 渗漏 (p =0.9878)。
综上所述,10 μM Nec-1可有效抑制TNPs的细胞毒性,且对细胞无毒性。
为了分析 Nec-1 的抗炎能力,将细胞与 TNPs 或 TNPs + Nec-1 一起培养。如图 4 所示,TNF-α (p =0.003), IL-1β (p =0.0013), IL-6 (p <0.0001) 和 IL-8 (p =0.0004)TNPs+Nec-1组显着低于TNPs组。
<图片>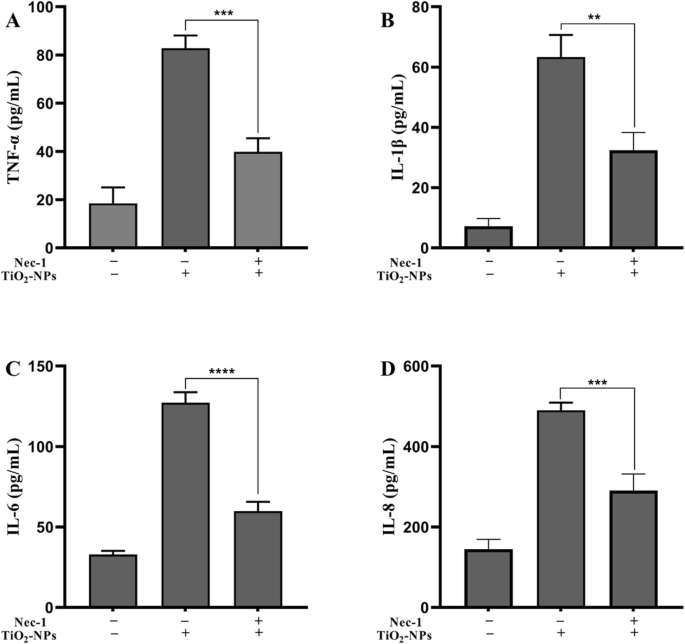
Nec-1 对 TNPs 暴露后炎症的影响。 ( ** p <0.01, *** p <0.001, **** p <0.0001)
这些结果表明Nec-1可以减轻TNPs暴露诱导的炎症反应。
沉默 RIP1 可减轻 TNP 引起的炎症损伤
为了确定 Nec-1 是否通过 RIP1 消除了 TNPs 诱导的炎症损伤,我们使用 siRNA 有效地沉默了细胞的 RIP1 表达(图 5,p <0.0001)。接下来,测量了突变体和野生型细胞上 TNPs 暴露后的炎症损伤。图 6a 表明 TNPs + si-RIP1 组的细胞活力显着高于 TNPs + si-NC 组(p =0.0002)。 LDH 生产(图 6b,p <0.0001) 和 TNF-α 水平 (p <0.0001), IL-1β (p =0.0001), IL-6 (p <0.0001) 和 IL-8 (p =0.0001) (图 7) TNPs + si-RIP1 组显着低于 TNPs + si-NC 组。这些结果表明TNPs通过RIP1促进炎症损伤。
<图片>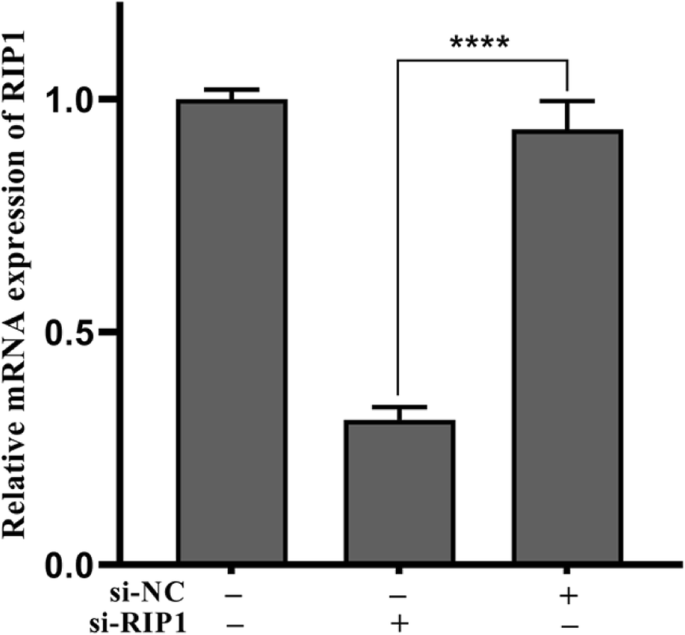
SH-SY5Y细胞转染si-RIP1后的mRNA表达。 ( **** p <0.0001)
<图片>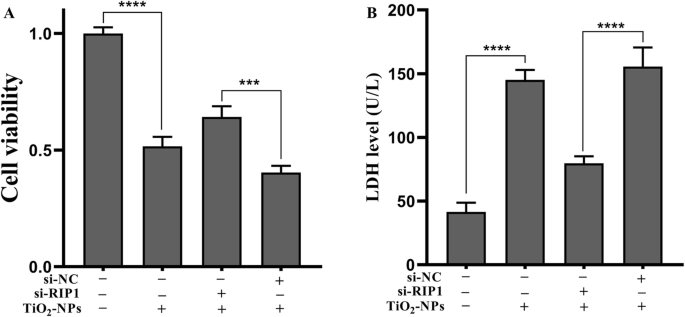
突变型和野生型细胞暴露于 TNPs 后的细胞活力和 LDH 泄漏。 ( *** p <0.001, **** p <0.0001)
<图片>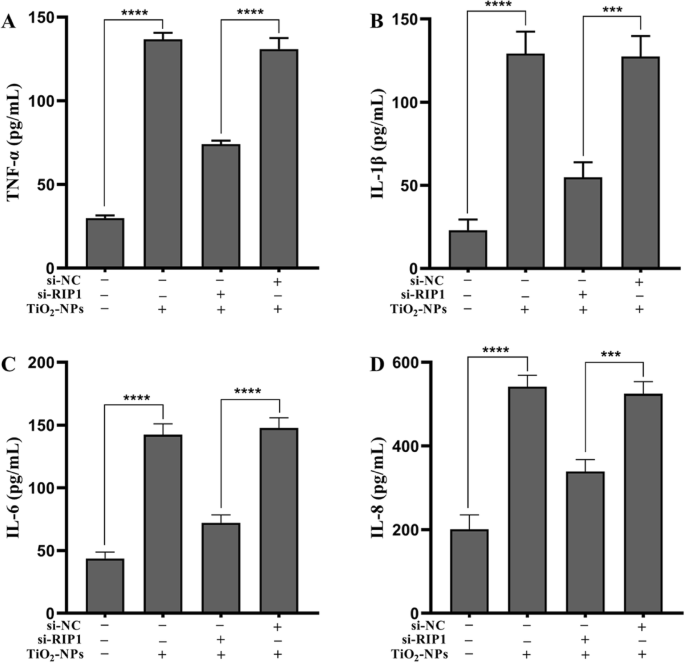
突变型和野生型细胞暴露于 TNP 后的炎症。 ( *** p <0.001, **** p <0.0001)
讨论
在这项研究中,我们发现 TNPs 暴露以剂量依赖性方式诱导对 SH-SY5Y 细胞的细胞毒性,并且坏死性凋亡与 TNPs 的神经毒性有关。为了探索坏死性凋亡的作用,我们将细胞暴露于含有或不含有 Nec-1(一种有效的坏死性凋亡抑制剂)的 TNP [17, 18],并测量了细胞活力、LDH 渗漏和炎性细胞因子的水平。我们的数据表明 Nec-1 联合治疗可以减轻 TNP 诱导的炎症损伤。由于研究表明 Nec-1 通过抑制 RIP1 活性发挥其作用,我们用 si-RIP1 转染细胞以确定 RIP1 是否参与了 Nec-1 的保护作用。用 TNPs 处理突变和野生型细胞,并评估炎症损伤。这表明在暴露于TNPs后,si-RIP1组的细胞活力高于si-NC组,并且si-RIP1组的LDH渗漏和炎性细胞因子水平低于si-NC组。 .总之,本研究首次揭示了坏死性凋亡参与了TNPs诱导的细胞炎症损伤。
我们在体外研究中再次确认了 TNP 的神经毒性。在 SH-SY5Y 细胞暴露于不同浓度的 TNP 不同孵育时间后,使用 CCK8 测定评估细胞活力。如图 1a 所示,TNPs 暴露以剂量依赖性方式降低细胞活力;暴露48小时和72小时后,与对照组相比,80和160 TNPs组的细胞活力显着降低(p <0.001)。为了进一步确认细胞毒性,我们还测量了 LDH 泄漏。图1b中的数据显示,暴露72小时后LDH的产生呈剂量依赖性增加,40、80和160组的LDH水平显着高于对照组(p <0.0001)。因为之前的研究表明,暴露于 TNP 可以促进神经炎症 [19],因此还测量了 TNF-α、IL-1β、IL-6 和 IL-8 的水平。图2说明与对照组相比,这四种炎性细胞因子的水平显着上调,并且在用20至160 μg/mL处理的组中显着更高(p <0.0001)。总之,我们的结果表明 TNP 可以以剂量依赖性方式诱导神经毒性,这与之前的研究一致。 TNP 可被神经元细胞吸收并抑制增殖 [20,21,22]。此外,TNPs暴露可能以剂量依赖性方式降低细胞活力并促进LDH渗漏[23,24,25,26,27]。
我们用 Nec-1 共同处理细胞以阐明坏死性凋亡是否与 TNP 引起的神经毒性有关。在细胞用指定浓度的 TNP 处理后,有或没有 Nec-1,细胞活力、LDH 渗漏和炎症被评估。图 3a 显示,与 10 μM Nec-1 共同处理显着抑制了 TNP 暴露后细胞活力的降低。同时,图 3b 显示用 10 μM Nec-1 处理没有增加 LDH 水平,并且用 TNPs + Nec-1 处理的细胞中 LDH 水平显着低于单独使用 TNP 组的细胞。接下来,我们测量了与 Nec-1 共同治疗对 TNP 诱导的炎症的影响。图 4 说明用 TNPs + Nec-1 处理的细胞中 TNF-α、IL-1β、IL-6 和 IL-8 的水平显着降低。这些结果再次验证了 Nec-1 可以保护细胞免受死亡和炎症过程的影响 [28, 29]。
最后,由于 RIP1 是 Nec-1 的靶点,我们评估了 RIP1 是否可以调节 TNP 的神经毒性。我们通过沉默 RIP1 表达构建了突变细胞(图 5)。图6和图7的数据显示,在TNPs暴露后,si-RIP1组细胞活力更高,LDH、TNF-α、IL-1β、IL-6和IL-8水平低于在 si-NC 中,这表明 TNPs 通过 RIP1 促进细胞毒性。同样,一些研究发现 RIP1 可以同时调节细胞死亡和炎症过程 [30, 31]。
尽管我们首次报道 Nec-1 可以通过抑制坏死性凋亡信号通路来减轻 TNP 的神经毒性,但我们的研究有一些局限性。首先,纳米尺寸材料的细胞毒性不同于它们的大块材料,并且可能在很大程度上受表面特性的影响 [32],这可以解释 Sebastián 等人发现的细胞活力增加的相反结果。 [33]。因此,有必要从细胞增殖、膜完整性、氧化应激、炎症、线粒体功能、细胞周期、细胞骨架和表观遗传学等多个方面综合评估纳米毒性对受试者的影响。其次,RIP1 可以招募 RIP3 形成功能性坏死体,这是 MLKL 的重要激活剂,以启动坏死性凋亡 [34]。我们假设 RIP3/MLKL 可能与 TNP 的神经毒性有关,这应该在未来的工作中进行探索。此外,还应评估 RIP3/MLKL 与 TNP 神经毒性的关联。第三,活性氧 (ROS) 作为 TNP 神经毒性的主要机制 [35, 36],据报道是 RIP1 的上游信号 [37]。因此,在TNPs的神经毒性中,ROS是否是RIP1的上游有待进一步讨论。第四,细胞应长期(超过 72 小时)暴露于无毒浓度(TNP <5 μg/mL)以评估慢性细胞毒性。第五,我们应该评估坏死性凋亡在更多神经元细胞系和原代人和动物神经元细胞中TNPs的神经毒性中的作用。
结论
我们的研究揭示了坏死性凋亡是另一种程序性细胞死亡机制,涉及 TNP 引起的神经毒性。在这项研究中,我们发现 TNP 可以以剂量依赖性方式诱导 SH-SY5Y 细胞的炎症损伤,而 Nec-1(一种有效的坏死性凋亡抑制剂)联合治疗可以减轻这些有害影响。 Nec-1 的靶点 RIP1 被 si-RNA 沉默,有效减轻了 TNPs 暴露引起的炎症损伤。总之,Nec-1通过靶向RIP1通路抑制TNPs暴露诱导的SH-SY5Y细胞的炎症损伤。 RIP1的上下游信号通路参与TNPs的神经毒性有待进一步评估。
材料和方法
TNP 制备和细胞培养
我们之前对 TNP 进行了表征,并根据我们之前发布的程序 [38] 制备了它们。简而言之,将 TNP 溶解在 RPMI 1640 中,浓度为 5、10、20、40、80 和 160 μg/mL。在室温下对 TNP 溶液进行灭菌和超声处理(300 W,10 分钟),以防止颗粒在处理前聚集。未暴露于 TNP 的培养基中的细胞作为对照组。 SH-SY5Y细胞购自中国科学院上海生命科学研究所细胞库,在添加10%胎牛血清(Gibco公司产品)的RPMI 1640培养基(HyClone,Logan,UT,USA)中培养。美国马萨诸塞州沃尔瑟姆的 Thermo Fisher Scientific 系列)、100 U/mL 青霉素和 100 μg/mL 链霉素,在 37 °C 下在含 5% CO2 的加湿培养箱中。
细胞活力和 LDH 渗漏检测
使用细胞计数试剂盒-8(CCK8.Dojindo,cat no.CK04)测量细胞活力。简而言之,1×10 4 将 SH-SY5Y 细胞置于 96 孔板中,并在暴露于 TNP 之前在 37°C(5% CO2)的培养箱中培养 24 小时。然后将细胞与不同浓度(0、1.25、2.5、5、10、20、40 和 80 μg/mL)的 TNP 再孵育 24 小时。处理后,将细胞与 CCK-8 再孵育 2 小时。接下来,通过将 96 孔板放入酶标仪(BioTek,Winooski,VT,USA)中,在 450 nm 处测量光密度(OD)。各组细胞活力,以百分比表示,计算公式为(OD处理-ODblank)/(OD对照-ODblank)×100%。
通过商业试剂盒(南京建成生物工程研究所,中国)测量膜完整性。 SH-SY5Y细胞暴露于不同浓度的TNPs 72 h后,根据生产商的说明分析LDH的分泌情况。
炎症反应
ELISA用于测量SH-SY5Y细胞产生的TNF-α、IL-1β、IL-6和IL-8的水平。简而言之,将SH-SY5Y细胞暴露于TNPs 72 h,并按照制造商的说明(Elabscience Biotechnology Co., Ltd.)收集上清液进行分析。
siRNA转染
按照制造商的方案,使用 Lipofectamine 2000 将 100 nM 浓度的 si-RIP1 或阴性对照 siRNA (si-NC) 转染到细胞中。通过实时 PCR 评估 siRNA 的基因沉默效率。
实时 PCR
根据制造商的说明,使用 RNAqueous 试剂盒(Ambion Inc.,Austin,TX,USA)分离暴露于不同浓度 TNP 的细胞的 RNA。 RIP1的相对表达通过实时PCR测量。 RIP1的相对mRNA水平被标准化为β-肌动蛋白表达。
统计分析
SPSS 11.0 软件(SPSS Inc., Chicago, IL, USA)用于数据分析。单因素方差分析用于比较组的平均值。 p <0.05 被认为具有统计学意义。
数据和材料的可用性
可从手稿中获得。
缩写
- TNP:
-
二氧化钛纳米粒子
- Nec-1:
-
Necrostatin-1
- RIP:
-
受体相互作用蛋白激酶
- MLKL:
-
混合谱系激酶结构域样蛋白
- ROS:
-
活性氧
纳米材料