

青蒿琥酯的纳米颗粒递送通过激活线粒体介导的细胞凋亡来增强抗肿瘤效率
摘要
青蒿素及其衍生物被认为具有广谱的抗癌活性,它们在肿瘤细胞中具有显着的抗癌作用。青蒿素及其衍生物吸收快,分布广,选择性杀伤肿瘤细胞。由于低浓度的青蒿琥酯主要依赖于溶瘤来诱导肿瘤细胞的细胞死亡,因此其抗肿瘤作用不理想且有限。为了获得更好的抗肿瘤效果,在本研究中,我们利用一种新的纳米技术设计了新型负载青蒿琥酯的牛血清白蛋白纳米颗粒,以实现青蒿琥酯的线粒体积累并诱导线粒体介导的细胞凋亡。结果表明,与游离青蒿琥酯依赖于溶胀性死亡相比,载有青蒿琥酯的牛血清白蛋白纳米颗粒显示出更高的细胞毒性,并且通过青蒿琥酯在线粒体中的分布诱导其显着的凋亡作用。该发现表明载有青蒿琥酯的牛血清白蛋白纳米粒通过上调细胞凋亡相关蛋白和促进细胞色素C的快速释放来破坏线粒体完整性并激活线粒体介导的细胞凋亡。
背景
青蒿素及其衍生物由于其抗疟活性高、毒性低而被广泛用于治疗疟疾。研究人员还发现,青蒿素及其衍生物具有显着的抗肿瘤活性,因为其毒副作用小,患者耐受性更高[1]。据报道,青蒿琥酯 (Ats) 确实抑制了肿瘤细胞的生长,并进一步在肿瘤细胞中诱导了显着的抗癌作用 [2,3,4]。一些实验表明,Ats在48小时后引起肿瘤细胞不同程度的凋亡和肿胀,并且细胞凋亡和肿胀的程度取决于Ats的剂量。在低浓度下,Ats 不会诱导肿瘤细胞发生明显的凋亡,并且 Ats 诱导的细胞死亡伴随着肿瘤样死亡 [5,6,7,8]。为了获得更大的抗肿瘤效果,应用了更高剂量的Ats,但这进一步证实了其严重的毒性和骨髓抑制。因此,有必要寻找有效的治疗方法,降低Ats的有效剂量,以提高其抗肿瘤效率[9,10,11]。发现线粒体在调节 Ats 的细胞凋亡和膨胀作用中起重要作用。线粒体还参与调节多种凋亡信号的转导过程 [12,13,14,15,16,17]。当线粒体受到药物攻击时,其通透性增强,膜电位降低,从而导致线粒体膜的子宫内膜肿胀,细胞色素C从线粒体中迅速释放到细胞质中[18,19,20]。此外,部分caspase家族蛋白被激活,诱导细胞凋亡的级联反应。
为了增强 Ats 的抗肿瘤作用,人们尝试了许多新技术来增加药物在肿瘤细胞中的分布或改善药物靶向递送到细胞器中以诱导细胞死亡 [21,22,23]。纳米粒子 (NPs) 作为靶向癌症治疗的关键工具已被广泛研究,并且它们已显示出巨大的潜力。由于纳米颗粒粒径小、表面积大,可以通过毛细血管进入血液循环,穿过内皮细胞间隙迁移到肿瘤部位,从而实现药物靶向分布,提高药物的生物利用度.此外,纳米颗粒可以通过生物材料长时间平稳地降解来控制药物的释放,最终延长消除半衰期,提高有效血药浓度,降低给药频率。最重要的是,载药纳米颗粒可以被递送到细胞内的特定位置,从而提高治疗效果[24,25,26]。
为了增强低浓度 Ats 的抗肿瘤作用,我们尝试设计新型负载 Ats 的牛血清白蛋白 (BSA) NP。由于肿瘤细胞内的低pH值,大量的氢质子积累存在于线粒体外膜或膜间隙中,相反,线粒体间膜因其化学成分和线粒体基质而富含负电荷分泌,这使得跨膜电位外部为正,内部为负,这可以有利于 BSA 的递送。然后,Ats 在线粒体中的大量积累可以有效地触发线粒体介导的细胞凋亡。结果表明,与游离 Ats 诱导的典型膨胀性死亡相比,Ats 在 BSA NPs 的介导下特异性转移到线粒体中,促进了线粒体介导的凋亡相关 caspase 蛋白的激活。这引发了显着的细胞凋亡,从而突出了更高的细胞毒性。
方法
材料
BSA 购自 Sigma-Aldrich 公司(美国密苏里州圣路易斯),Ats 购自桂林制药公司(中华人民共和国桂林)。 SMMC-7721细胞和Plc细胞购自中国科学院生物化学与细胞生物学研究所(中华人民共和国上海)。所有其他购买的化学品均为分析级;它们是从各种供应商处获得的。
加载 Ats 的 BSA NP 的制备和表征
根据先前报道的文献 [27],通过去溶剂化方法制备了负载 Ats 的 BSA NP。简而言之,通过在 37°C 下将 1.0 mL 含有一定量 Ats 的无水酒精快速滴入 0.5 mL BSA 溶液中直至乳白色,制备负载 Ats 的 BSA NP。通过旋转蒸发去除乙醇,载有 Ats 的 BSA NPs 进一步从培养基中沉淀出来,然后加入 8% 戊二醛水溶液(0.5 μL/mg BSA)以在悬浮液搅拌下诱导粒子交联一段时间24 小时。最后,使用 Brookhaven Zetasizer(Brookhaven Instruments Corporation, Holtsville, NY, USA)收集纳米颗粒并用去离子水洗涤 3 次以进一步分析它们的物理特性,包括它们的流体动力学直径、多分散指数 (PDI)、zeta 电位和形态和透射电子显微镜(JEM-1200EX;JEOL,东京,日本)。使用先前报道的方法[27]估计了Ats在BSA NPs中的包封效率的测定。
MTT 分析
两种肿瘤细胞系 SMMC-7721 细胞和 Plc 细胞分别与 20% 胎牛血清 (FBS) 孵育。细胞生长密度调整为l × l0 6 细胞计数/mL,然后将细胞悬液稀释至l × l0 5 细胞/毫升。将稀释的悬浮液进一步单独加入96孔板中(每孔100 μL,约1 × 10 4 细胞/孔)在 5% CO2 和 95% O2 的条件下,在 37°C 下连续孵育 24 小时。在存在具有不同浓度 Ats 的游离 Ats 或负载有 Ats 的 BSA NP 的情况下,将培养基替换为无血清培养基,然后孵育 24 小时。将总共 50 μL 的 3-(4,5-二甲基噻唑-2-基)-2,5-二苯基溴化四唑 (MTT) (5 mg/mL) 添加到每个孔中并孵育 4 小时以终止培养。当四唑鎓染料 MTT 被还原为不溶性甲臜时,96 孔板以 1000 rpm 离心 5 分钟,从每孔中倒出上清液,然后加入 150 μL 的二甲基亚砜 (DMSO),完全溶解了晶体。使用酶标仪(Syneray-2;BioTek Instruments, Inc., Winooski, VT, USA)在 490 nm 处测量溶液的吸光度。
细胞内 BSA NP 组的细胞内分布
选择对数期的SMMC-7721细胞和Plc细胞进行胰蛋白酶消化处理;细胞浓度调整为l × l0 6 细胞/毫升。接下来,将培养的细胞加入6孔细胞培养板进行贴壁,除去培养基,加入罗丹明B标记的BSA NPs。细胞核在 37°C 下用 Hoechst(蓝色)染色 15 分钟,线粒体用 Mitotracker Green FM 染色。使用共聚焦激光扫描显微镜(FluoView FV10i;Olympus Corporation,Tokyo,Japan)追踪细胞内BSA NPs的位置。
线粒体膜电位变化
JC-1 可用于确定线粒体膜电位的变化。当线粒体膜电位高时,JC-1能够自由穿过细胞膜并在线粒体内形成聚集体,呈现红色荧光(激发波长525nm;发射波长590nm);当线粒体膜电位降低时,JC-1 从线粒体基质转移到细胞质中,形成绿色荧光单体(激发波长,490 nm;发射波长,530 nm)。 SMMC-7721细胞和Plc细胞分别接种于共聚焦培养皿中,密度为l × l0 6 细胞/mL,用于连续孵育 12 小时。接下来,弃去培养基,将含有 Ats 或负载有 Ats 的 BSA NP 的分散体的无血清培养基加入培养皿中。 9 h后弃去培养基,用PBS洗涤细胞两次,然后加入2 mL浓度为2 μmol/L的JC-1;然后将细胞在黑暗条件下在 37°C 下孵育 30 分钟。采用激光扫描共聚焦显微镜(FluoView FV10i;Olympus Corporation)观察线粒体膜成像变化。
内质网 (ER) 的 ROS 产生测量和染色
细胞用20% FBS培养,细胞生长密度调整为l × l0 6 细胞数/mL,按细胞计数;然后将细胞悬液稀释至l × l0 5 细胞/毫升。将稀释的悬浮液进一步加入 96 孔板中(每孔 100 μL,约 1 × 10 4 细胞/孔)以在 37°C 下在 5% CO2 和 95% O2 下连续孵育 24 小时。其次,将游离的 Ats 和负载有 Ats 的 BSA NP 与细胞一起孵育 6、12 和 24 小时,然后与 10 μM 的 2,7-二氯荧光素二乙酸酯(DCFH-DA;Sigma-Aldrich Co.)连续孵育大约 30 分钟。使用冰冷的 PBS 缓冲液清洗细胞 3 次以去除未内化的 NPs。使用酶标仪(Synergy-2;BioTek Instruments)检测细胞内 DCF 荧光强度,在 485 nm 激发并在 530 nm 发射,以研究氧化应激的程度。测试组用 SMMC-7721 细胞和 Plc 细胞处理 24 小时,然后将 ER-Tracker Blue-White DPX 探针(Molecular Probes,Eugene,OR,USA)加入细胞中孵育 30 分钟。弃上样液,PBS洗涤细胞后,共聚焦激光扫描显微镜观察ER的形态变化。
流式细胞术评估细胞肿瘤和凋亡
根据我们之前研究的方案 [28],使用膜联蛋白 V-异硫氰酸荧光素 (FITC)/碘化丙啶 (PI) 染色测定来评估游离 Ats 和负载 Ats 的 BSA NP 诱导的细胞坏死和凋亡。细胞用胰蛋白酶裂解并以l × l0 6 的浓度接种到六孔板中 细胞/mL 连续孵育 24 小时。接下来,去除培养基并将含有游离 Ats 和负载 Ats 的 BSA NPs 的无血清培养基加入孔中。处理后,收集细胞并悬浮在含有 PI 和 FITC 标记的膜联蛋白 V(AV-FITC)的 Nicoletti 缓冲液(北京 4A 生物技术有限公司,北京,中华人民共和国)中。共聚焦激光扫描显微镜观察细胞形态变化。为了验证负载Ats的NPs诱导的细胞凋亡和膨胀率,通过流式细胞术定量了早期凋亡(Q4)、膨胀(Q2)、坏死(Q1)和活细胞(Q3)的百分比。
细胞凋亡相关蛋白和细胞色素 C 的蛋白质印迹分析
当游离的 Ats 或负载有 Ats 的 NPs 与 SMMC-7721 细胞孵育 24 小时时,进行了蛋白质印迹分析以确定相关蛋白质的水平。用冰冷的放射免疫沉淀试验 (RIPA) 缓冲液裂解细胞,该缓冲液含有蛋白酶抑制剂混合物和磷酸酶抑制剂(Roche,Basel,Switzerland)。使用改良的 BSA 检测试剂盒(Thermo Fisher Scientific,Waltham,MA,USA)测定蛋白质浓度,并在加载到 10% 十二烷基硫酸钠 (SDS)-聚丙烯酰胺凝胶电泳 (PAGE) 之前进行标准化。使用UVP凝胶分析系统(iBox Scientia 600; UVP, LLC., Upland, CA, USA)对靶蛋白的水平进行拍照和分析。
结果
加载 Ats 的 BSA NP 的特征和细胞活力研究
在图 1a、b 中观察到加载 Ats 的 BSA NP 呈球形,并且它们均匀分散,PDI 较低,为 0.016。负载 Ats 的 BSA NP 的平均粒径约为 99.9 ± 2.3 nm,zeta 电位为负值,约为 –25.6 ± 4.3 mV。释放曲线显示出平滑和持续的释放,如图 1c 所示。与体外培养基中游离 Ats 的快速释放相比,由于 BSA 的不断降解,被包裹在 BSA NPs 核心的 Ats 从 NPs 内部缓慢扩散到培养基中并显示出平稳和持续的释放模式。超过 85% 的游离 Ats 在前 6 小时内完全释放,而在 48 小时内从 NPs 释放到介质中的药物累积总量为 78.9%。这表明纳米粒可以通过生物材料长时间平稳的降解来控制药物的释放,从而延长消除半衰期,提高有效血药浓度,降低给药频率。
<图片>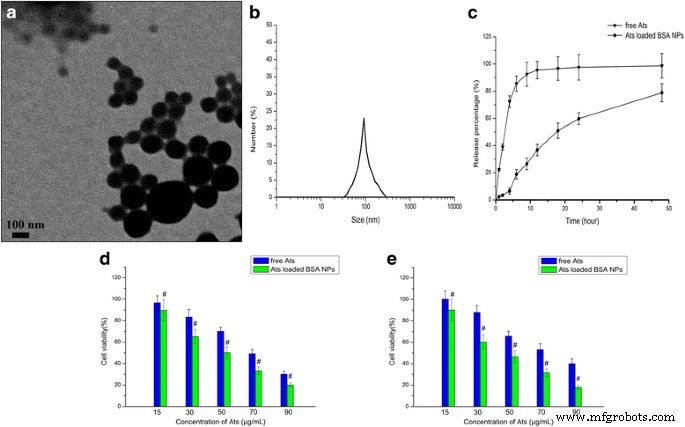
加载 Ats 的 BSA NP 的表征。 一 加载 Ats 的 BSA NP 的 TEM 图像。 b 获得的加载了 Ats 的 BSA NP 的动态光散射 (DLS) 分析。 c 加载 Ats 的 BSA NP 在 37°C 下 pH 为 7.4 的磷酸盐缓冲盐水中 48 小时的体外释放曲线。 SMMC-7721 细胞的活力 (d ) 和 Plc 单元 (e ) 与不同量的游离 Ats 和负载 Ats 的 BSA NP 孵育 24 小时。数据表示为平均值 ± SD (n =3)。 # P <0.05 相对于相应的游离 Ats
MTT 用于检测游离 Ats 和负载 Ats 的 BSA NPs 在不同时间间隔对 SMMC-7721 细胞和 Plc 细胞的抑制作用。结果(图1d,e)表明游离Ats的细胞毒性随着药物浓度的增加而增加,并且负载Ats的BSA NPs显示出逐渐增强的细胞毒性。这证明了 Ats 和载有 Ats 的 BSA NPs 抑制了肿瘤细胞的生长,并且抑制率取决于 Ats 的剂量。与游离的 Ats 相比,载有 Ats 的 BSA NPs 在两种细胞中都表现出更高的细胞毒性和更高的敏感性,并且它们导致更大的细胞抑制。如图 1d、e 所示,与游离 Ats 相比,用负载 Ats 的 BSA NPs 处理两种细胞导致 24 小时细胞活力显着降低。用负载有 Ats 的 BSA NPs 处理的 SMMC-7721 细胞和 Plc 细胞的 50% 最大抑制浓度 (IC50) 值在 24 小时分别为 50.1 和 44.9 μg/mL,与获得的值 69.2 和 74.9 进行比较在用游离 Ats 处理的细胞中,24 小时时为 μg/mL。这表明当Ats被加载到BSA NPs中时,它可能会在NPs的介导下改变其在细胞内的位置,最终杀死更多的细胞。
BSA NPs 的体外细胞摄取
通过共聚焦激光扫描显微镜观察两种类型肿瘤细胞中BSA NPs的细胞内分布和位置,如图2所示。罗丹明B标记的NPs与细胞共培养3小时后,红色荧光清晰可见在细胞质中;随着时间的推移,大多数 BSA NPs 被细胞内化并扩散到细胞质中,显示出增强的时间依赖性红色荧光。还观察到位于细胞质中的 BSA NPs 与线粒体位于同一位置,黄色荧光的出现证明了这一点,这表明罗丹明 B 标记的 NPs 固有的红色荧光和由线粒体指示剂 MitoTracker® green FM 已合并。这证明内化的BSA NPs可以在线粒体内特异性积累,突出了在BSA NPs的介导下将Ats递送至线粒体的可能性。
<图片>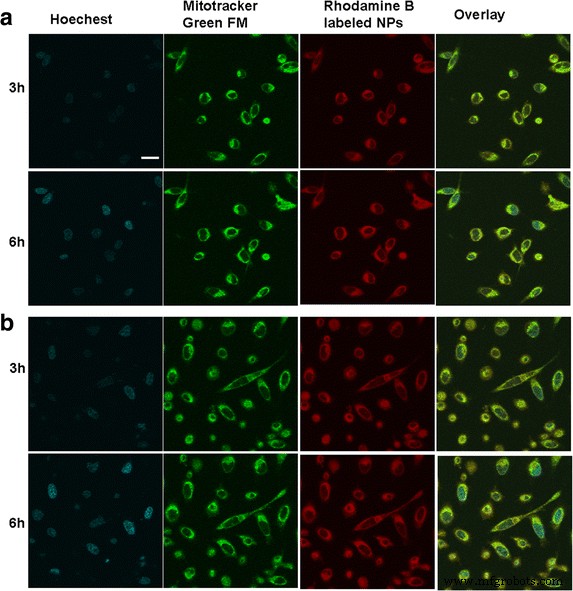
BSA NPs 与不同肿瘤细胞孵育后的体外细胞分布。 SMMC-7721 细胞的荧光图像 (a ) 和 Plc 单元格 (b )。 比例尺 , 100 μm
线粒体膜电位分析
为了阐明在线粒体中递送 Ats 后加载 Ats 的 BSA NP 是否干扰线粒体功能,确定了线粒体膜电位的变化。图 3 表明,在 JC-1 染色后,用游离 Ats 处理的肿瘤细胞中的大部分线粒体表现出强红色荧光和弱绿色荧光强度。这表明大部分 JC-1 以聚集状态存在,增强了线粒体膜的完整性和更高的潜力。相反,当 JC-1 染色负载 Ats 的 BSA NPs 处理的细胞时,两种肿瘤细胞中的线粒体都表现出更强的绿色荧光,表明线粒体膜受到严重破坏,其电位显着降低。综上所述,证明Ats在BSA NPs的介导下成功递送至线粒体,导致线粒体膜去极化。
<图片>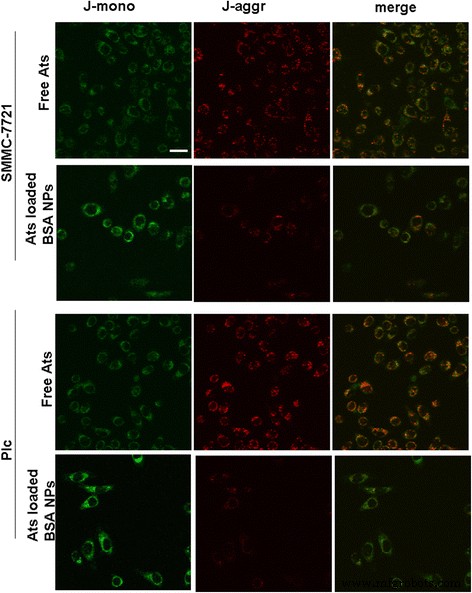
将游离 Ats 和负载有 Ats 的 BSA NPs 与 SMMC-7721 细胞和 Plc 细胞孵育后,线粒体膜电位的成像变化。 比例尺 , 100 μm
ER 的 ROS 产生测量和染色
广泛证实大量ROS的产生可引起线粒体内膜磷脂过氧化,还可诱导线粒体膜电位降低,从而导致细胞色素C的快速释放。我们使用DCFH- DA作为荧光探针检测ROS的变化。 DCFH-DA自由通过细胞膜进入细胞,经酯酶水解转化为DCFH。生成的DCFH不能通过细胞膜,很容易加载到细胞中。细胞内 ROS 将非荧光 DCFH 氧化为具有绿色荧光颜色的 DCF。因此,DCF荧光检测可以指示细胞内ROS的水平。
当两种细胞都用游离的 Ats 和载有 Ats 的 BSA NPs 处理一段时间后,细胞内 ROS 的量也有所增加,呈现出时间依赖性。与游离的 Ats 相比,负载 Ats 的 BSA NPs 处理的 SMMC-7721 细胞和 Plc 细胞中 ROS 的产生显着增强。图 4a 表明,与经处理的 SMMC-7721 细胞和 Plc 细胞相比,暴露于加载了 Ats 的 BSA NP 48 小时的 SMMC-7721 细胞和 Plc 细胞中的 ROS 水平已分别增加至 1.53 倍和 1.28 倍有免费的 Ats。这支持了 NPs 加速细胞内 ROS 产生的观点。与对照组和游离 Ats 相比,在用负载 Ats 的 BSA NPs 处理后,ER-Tracker Blue-White DPX 作为 ER 特异性染料的荧光染色强度显着增加,表明也触发了 ER 应激在负载 Ats 的 NPs 处理的细胞中,ROS 水平相应增加。这一发现强调了 Ats 特异地位于线粒体中,由 BSA NPs 介导;这导致细胞内氧自由基水平显着增加,从而触发ER应激的诱导并激活线粒体途径以诱导caspase依赖性细胞凋亡。
<图片>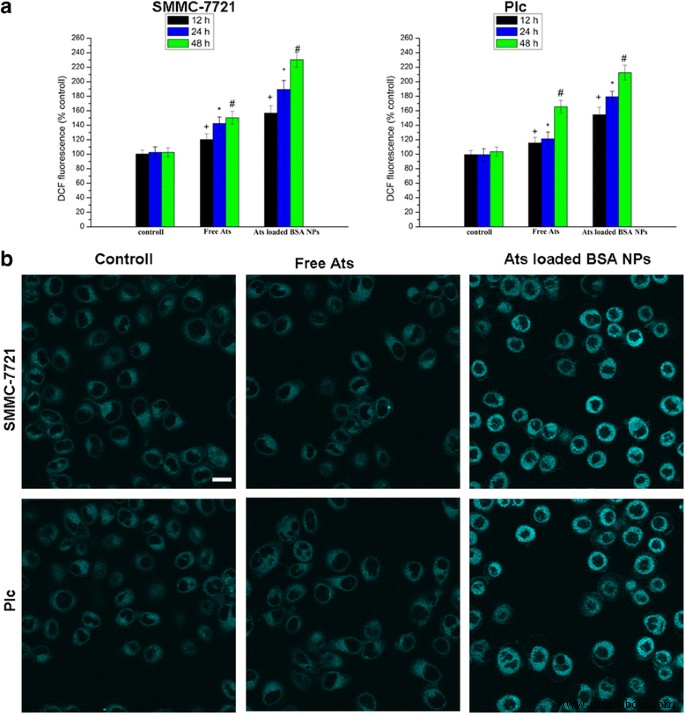
在不同时间用游离 Ats 和负载 Ats 的 BSA NPs 处理的细胞中 ROS 生成的量化 )。 ER 染色与 ER-Tracker 蓝白 DPX 探针 (b )。 比例尺 , 100 微米。数据表示为平均值 ± SD (n =3)。 + P <0.05 与对照组在 12 小时相比,*P <0.05 与对照组在 24 小时相比, # P <0.05 vs 24 h 对照组
细胞凋亡和坏死的评估
通过膜联蛋白 V-FITC/PI 染色测定处理细胞。活细胞不与膜联蛋白 V-FITC/PI 结合,因此不出现荧光。凋亡细胞不与 PI 结合,但它们被 Annexin V-FITC 染色,产生绿色荧光。相反,对于膨胀细胞,它们的细胞膜受到了一定程度的破坏,细胞核被扩张分解成碎片,从而同时显示出绿色和红色荧光。如图 5a 所示,与对照组相比,当游离 Ats 和负载 Ats 的 NPs 与细胞孵育 24 h 时,细胞中观察到强烈的绿色和红色荧光,表明游离的 Ats 和负载 Ats 的 BSA NP诱导肿瘤细胞浸润和凋亡。尤其是负载Ats的BSA NPs处理后,Annexin V-FITC和PI获得的染色荧光强度显着增加,表明负载Ats的NPs处理的细胞的肿胀和凋亡程度显着增强。
<图片>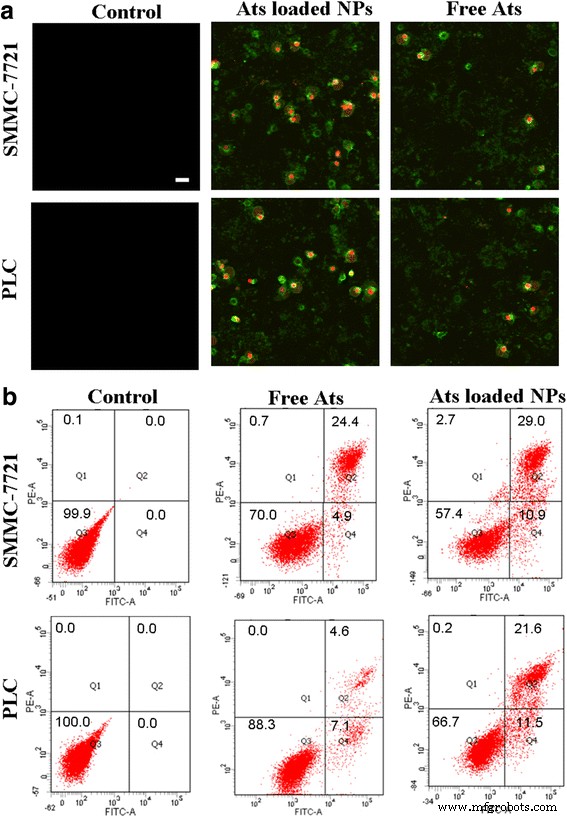
使用膜联蛋白 V-FITC/PI 染色测定用游离 Ats 和负载 Ats 的 BSA NPs 处理的细胞的超微结构变化的形态学 (a )。 比例尺 , 100 微米。分别与游离 Ats 和负载 Ats 的 BSA NPs 孵育 24 小时后,流式细胞仪分析细胞凋亡和肿块 (b )
图 5b 显示了早期凋亡 (Q4)、膨胀 (Q2)、坏死 (Q1) 和活细胞 (Q3) 的百分比。这一发现表明,当细胞用游离 Ats 处理时,膨胀率逐渐增加到 24.4% 和 4.6%,而 SMMC-7721 细胞和 Plc 细胞的凋亡率分别保持在 4.9% 和 7.1%,表明游离 Ats 触发发生溶瘤和细胞凋亡导致细胞死亡。相反,加载了 Ats 的 BSA NPs 显着提高了细胞凋亡和肿瘤的发生率。 SMMC-7721 细胞的凋亡率显着增加到 10.9%,Plc 细胞增加到 11.5%。在 SMMC-7721 细胞中,膨胀率增加到 29.0%,在 Plc 细胞中增加到 21.6%。这表明在 BSA NPs 的介导下,Ats 的线粒体递送通过增强肿瘤细胞和凋亡作用加速了肿瘤细胞的死亡。负载Ats的BSA NPs触发细胞凋亡信号转导过程,促进线粒体介导的细胞凋亡级联反应。
Western Blot 分析
为了探索细胞死亡对游离 Ats 和负载 Ats 的 NPs 诱导的细胞凋亡的依赖性,进行了蛋白质印迹分析以检测细胞凋亡蛋白的表达。发现在加载了 Ats 的 NPs 处理的 SMMC-7721 细胞中,Bax 蛋白的细胞内表达水平显着增加(图 6)。这一发现表明,在 BSA NPs 的帮助下,Ats 在线粒体中积累并导致线粒体功能障碍。细胞质中的Bax单体蛋白被转移到线粒体外膜并进行寡聚化,在线粒体外膜中形成蛋白质通道,从而进一步导致膜通透性增加。细胞质中细胞色素C的表达水平也特别显着增强,caspase-3和caspase-9的表达呈上升趋势。因此,由于线粒体较高的膜通透性,细胞色素C迅速释放到细胞质中,激活细胞死亡信号蛋白(caspases),促进细胞凋亡的级联反应。相比之下,游离Ats对凋亡相关蛋白和细胞色素C的表达没有显着差异,表明游离Ats不会触发线粒体介导的细胞凋亡,主要依赖于细胞凋亡导致细胞死亡。如我们的蛋白质印迹分析所示,BSA NPs 增强了药物在线粒体中的积累并激活了线粒体介导的细胞凋亡作用,从而导致显着的细胞凋亡并增加了主要细胞凋亡相关蛋白的表达。
<图片>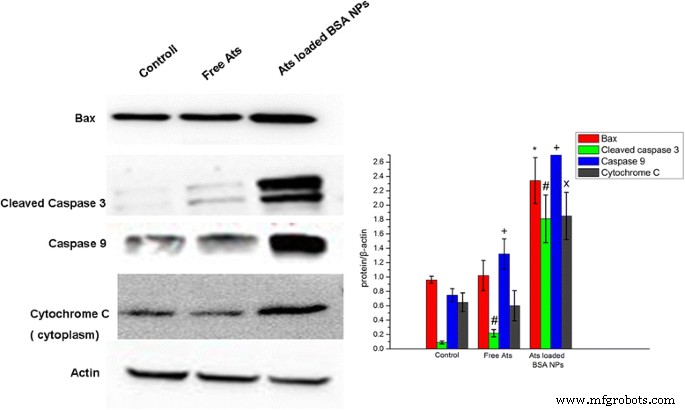
SMMC-7721 细胞中裂解的 caspase-3、caspase-9、Bax 和细胞色素 C 的表达水平的蛋白质印迹分析。 *P <0.05 相对于对照组的Bax蛋白表达; # P <0.05 与对照组裂解的 caspase-3 表达相比; + P <0.05与对照组的caspase-9蛋白表达相比; x P <0.05 与对照组的细胞色素 C 蛋白表达相比。数据表示为平均值 ± SD (n =3)
讨论
肿瘤和细胞凋亡代表细胞死亡的两种不同方式。细胞凋亡是发生在多细胞生物中的程序性细胞死亡的活跃过程。另一方面,Oncosis 描述了一种不依赖于半胱天冬酶的细胞死亡,其特征是肿胀、通透性增加和膜破裂,这通常被称为坏死。这种形式的细胞死亡被认为是意外和不受控制的。根据我们的调查,我们发现 Ats 抑制了肿瘤细胞的生长,并且抑制率取决于 Ats 的剂量。 Ats 主要取决于肿瘤的程度并导致细胞死亡;它还以溶瘤的形式激活不依赖于半胱天冬酶的细胞死亡。相反,与明显的肿瘤样死亡的发生分开,当肿瘤细胞用加载了 Ats 的 BSA NPs 处理时,加载了 Ats 的 BSA NPs 被内化到细胞质中并迅速定位在线粒体内以释放 Ats,这是由NP。线粒体中的 Ats 产生 ROS 并引发内质网应激;它通过降低线粒体膜电位、释放细胞色素 C 和促进 Bax、裂解的 caspase 3 和 caspase 9 的蛋白质表达,进一步激活了线粒体介导的 caspase 依赖性细胞凋亡途径。线粒体递送Ats并增强肿瘤和细胞凋亡的程度以诱导细胞死亡,从而增加药物的细胞毒性并诱导显着的细胞死亡。
结论
简而言之,我们澄清了肿瘤细胞中的游离Ats强烈依赖于肿瘤的程度,以抑制肿瘤细胞以类肿瘤死亡的形式增殖;因此,该药物的细胞毒性是有限的并且是不可取的。相比之下,加载了 Ats 的 BSA NPs 激活了线粒体凋亡途径并同时触发了膨胀效应;它们共同增强了 Ats 的协同抗肿瘤功效。这项研究的结果强调了负载 Ats 的 BSA NPs 在增强 Ats 的细胞毒性和凋亡作用方面的重要性,并且它们进一步表明了 BSA NPs 在使 Ats 诱导的细胞死亡途径多样化中的作用。 Compared with free Ats, Ats-loaded BSA NPs induced greater cytotoxicity and significant cell apoptosis effects in tumor cells.
纳米材料
- 共发射极放大器
- COVID-19 对制造业的影响
- 将活性氧种类的毒性调整到高级肿瘤治疗中
- 环境响应金属-有机框架作为肿瘤治疗的给药系统
- Li/Nb 比对 Li-Nb-O 化合物制备和光催化性能的影响
- DNA 四面体递送增强多柔比星诱导的 HT-29 结肠癌细胞凋亡
- 超材料中表面等离子体激元和磁偶极子共振的耦合效应
- 背面有黑硅层的晶体硅太阳能电池的研究
- 评估由食用菌根真菌 Tricholoma crassum 合成的蛋白质包覆金纳米粒子的抗菌、凋亡和癌细胞基因传递特性
- Thiacalix[4] 芳烃消除锌阳离子对肌球蛋白 ATP 酶活性的抑制作用
- 提高柔性纤维染料敏化太阳能电池的光电转换效率
- 农业自动化的积极影响