

金纳米粒子对人类肝脏微粒体中睾酮代谢的影响
摘要
金纳米颗粒 (AuNP)-蛋白质电晕复合物可以通过改变其理化特性来改变细胞色素 P450 (CYP) 介导的睾酮 (TST) 代谢。我们研究了 NP 大小、表面化学和蛋白质电晕对混合人肝微粒体 (pHLM) 中 TST 代谢的影响,采用 40 和 80 nm AuNP 以支化聚乙烯亚胺 (BPEI)、硫辛酸 (LA) 和聚乙二醇 (PEG) 功能化) 以及人血浆蛋白电晕 (PC)。 AuNP 介导的 TST 代谢的个体差异也在包含不同 CYP 活性水平的单一供体 HLM 中表征。 40 nm AuNP 和较小程度的 80 nm AuNP 对 pHLM 中总共五种 TST 羟基化代谢物的产生产生抑制作用,但 PC 减轻了它们。同时,裸 AuNP 增加了雄烯二酮的产量。 TST 代谢的个体间变异发生在单个供体 HLM 中。在大多数情况下,40 和 80 nm 裸露和 PC AuNP 在非抑制浓度下基本上抑制了 TST 代谢,但 PC PEG-AuNP 增加了雄烯二酮。这些研究有助于更好地理解 AuNP 通过改变 TST 代谢作为 TST 干扰物的作用,并可用于筛选其他 NP 作为潜在的内分泌干扰物。
介绍
由于其独特的光学和物理特性,金纳米粒子 (AuNP) 已广泛用于药物输送、医学诊断和癌症治疗以及消费品,即化妆品、食品包装 [1,2,3]。暴露于蛋白质混合物后,NP 与蛋白质结合并形成蛋白质电晕,这改变了表面化学、吸附的蛋白质构象和随后的生物反应,即 NP 毒性、细胞 NP 摄取、细胞色素 P450(CYP)的催化活性) 酶对药物 [4,5,6,7]。对原代上皮细胞和癌细胞系的体外研究表明,AuNP 对人肝细胞、肝癌细胞系 C3A 和精子细胞具有毒性 [6,7,8]。但是在 NP 周围形成的蛋白质电晕以表面化学依赖性方式强烈减弱或增强 AuNP 的毒性 [6, 7]。蛋白质电晕干扰了人肝细胞、肾近端小管细胞、HepG2 细胞、C3A 细胞系中 AuNP 的细胞摄取,无论它们的大小和表面电荷如何[6, 7, 9,10,11,12]。
肝 CYP 酶主要参与内源性和外源性化合物的合成和/或代谢,但范围广泛的药剂,即药物、杀虫剂或 NP,会反向影响类固醇激素的合成、代谢和/或解毒,从而产生药理作用和生理功能 [13,14,15,16,17]。睾酮 (TST) 是一种重要的雄激素和 CYP3A4 特异性底物(主要转化为 6β-OH TST),以区域和立体选择性方式 [18]。在 I 期代谢期间,TST 也被 CYP3A4 羟基化为 2β-OH TST,并被 CYP2D6 脱烷基化为雄烯二酮 (AD) [17, 19]。对人肝细胞、C3A 细胞系、人肝微粒体 (HLM) 和重组 CYP 酶的体外研究表明,裸露和蛋白冠包覆的 AuNP 可调节多种 CYP 酶,包括 CYP1A2、2C9、2C19、2D6、2E1、和 3A4 [6, 7, 20, 21]。其他金属 NP,裸 AgNP 也抑制 CYP3A4 介导的 HLM 中 6β-OH TST 的产生 [22]。用支链聚乙烯亚胺 (BPEI) 和硫辛酸 (LA) 功能化的 AuNP 降低了 C3A 细胞系中 CYP3A4 的活性,但人血浆蛋白冠 (PC) 会减弱它 [7]。相比之下,裸(无 PC)和 PC BPEI-AuNP 对人肝细胞中的 CYP2C9 和 3A4 具有抑制作用,而与 NP 大小无关 [6]。
体内研究报告称,小尺寸的 AuNP(4 和 13 nm)主要在雄性 BALB/c 小鼠的肝脏和脾脏中积累,并诱导肝脏 Cyp1a1 和 2b 基因的表达 [23]。其他金属纳米颗粒、氧化锌纳米颗粒对雄性 Sprague Dawley 大鼠肝脏 CYP1A2、2C11 和 3A2 活性有抑制作用,肝脏病理变化增加 [24]。
迄今为止,在不存在和/或存在生物相关蛋白电晕的情况下,AuNP 如何与 CYP 介导的 TST 代谢(TST 的羟基化和脱烷基化)相关联,我们知之甚少。本研究的目的是研究 PC 对 40 和 80 nm 阳离子 BPEI AuNP、阴离子 LA AuNP 和中性聚乙二醇 (PEG) AuNP 理化性质的影响。 AuNP 对有和没有 PC 的 CYP 介导的 TST 代谢的影响将使用 pHLM 进行表征。还将研究含有不同程度CYP酶的单供体HLM内TST代谢的个体差异。
方法/实验
化学品
2,3,4- 13 C3 睾酮 (CAS#327048-83-9) 和 6β-羟基睾酮 (6β-OH TST, CAS#62-99-7) 从 MilliporeSigma (St. Louis, MO) 获得。睾酮 (TST, CAS# 58-22-0), 2α-羟基睾酮 (2α-OH TST, CAS#004075-14-3), 2β-羟基睾酮 (2β-OH TST, CAS#10390-14-4), 6α -羟基睾酮(6α-OH TST,CAS#2944-87-8),11β-羟基睾酮(11β-OH TST,CAS#1816-85-9),15β-羟基睾酮(15β-OH TST,CAS#39605-73- 7), 16α-羟基睾酮 (16α-OH TST CAS#63-01-4), 16β-羟基睾酮 (16β-OH TST, CAS#17528-90-4), 11-酮睾酮 (CAS#564-35-2) 、雄烯二酮 (AD, CAS#63-05-8)、4-羟基雄烯二酮 (CAS#566-48-3) 和 11β-羟基雄烯二酮 (CAS#382-44-5) 购自 Steraloids (Newport, RI )。 LC-MS 级乙腈和甲酸购自 Fisher Scientific(Fair Lawn,新泽西州),而超纯水由 Merck KGaA(德国达姆施塔特)的 Synergy® UV-R 系统内部生产。如未注明,所有其他试剂均购自 MilliporeSigma (St. Louis, MO)。
人肝微粒体
汇集的人肝微粒体 (pHLM)(200 名供体,100 名男性和 100 名女性)和单个供体肝微粒体获自康宁公司(北卡罗来纳州夏洛特)。 pHLM 由供应商汇集,而不是由单个供体 HLM 汇集而成。本研究中使用的单一供体 HLM 的特征和选定的细胞色素 P450 (CYP) 酶活性见附加文件 1:表 S1。
金纳米粒子合成
用阳离子支化聚乙烯亚胺 (BPEI)、阴离子硫辛酸 (LA) 和中性聚乙二醇 (PEG) 功能化的 Biopure™ 40 和 80 nm 球形 AuNP 购自 nanoComposix (San Diego, CA)。通过在碳酸钾水溶液中还原四氯金酸氢(III)水合物(HAuCl4·3H2O)合成核心材料,并进行老化过程和切向流过滤(TFF)。通过添加二氢硫辛酸 (0.2:1, w /w ) 或巯基甲氧基封端的 PEG (Laysan Bio Inc., Arab, AL) (0.5:1, w /w ),分别用 TFF 洗涤和无菌过滤。通过将 LA 的羧酸与 BPEI 的胺连接起来,通过 EDC/NHS 化学合成了 BPEI 功能化的表面。通过 TFF 洗涤和随后的离心去除未结合的 BPEI。
人血浆蛋白冠的制备
混合人血浆(HP,n =5) 从 Biological Specialty Corp. (Colmar, PA) 获得。 40 和 80 nm AuNP 与人血浆一起孵育,生理血浆体积占总血容量的 55% (v /v ) 在 37°C、250 rpm 的轨道摇动/旋转培养箱中培养 1 小时。孵育结束时,通过 20,000×g 离心收集 NP 周围的人血浆蛋白电晕 (PC) 在 20°C 下 20 分钟,然后进行三次磷酸盐缓冲盐水 (PBS) 洗涤。通过离心丢弃未结合和松散结合的蛋白质。所得PC AuNP分散在PBS中,用于表征理化性质及其与TST的相互作用。
AuNP 的物理表征
通过动态光散射 (DLS) 和透射电子显微镜 (TEM) 测量粒度和表面特性。使用 Zetasizer Nano-Zs 测量去离子 (DI) 水中的 40 和 80 nm 裸露(无 PC)BPEI-、LA-和 PEG-AuNP 以及 PBS 中的 PC AuNP 的流体动力学直径 (DH) 和 zeta 电位(Malvern Instruments, Worcestershire, UK) 25 °C 下 0 小时。在微粒体孵育缓冲液 (pH 7.4) 中,在 37°C 下 0 分钟和 45 分钟,还获得了裸露和 PC AuNP 的 DH、多分散性指数 (PDI) 和 zeta 电位。样品测量五次,每次 10 秒,共 11 个子运行。 TEM 表征了裸金纳米粒子和 PC 金纳米粒子的形态。所有 AuNP 都放置在涂有 formvar 的铜网上,并在 Tecnai G2 Spirit BioTWIN 上以 120 kV 的加速电压和牛津探测器(FEI Company, Hillsboro, OR)观察。 GATAN 显微镜套件(GATAN Inc., Pleasanton, CA)测量 AuNP 直径。使用 Spectra Max i3 多模式酶标仪(Molecular Devices, Sunnyvale, CA)测量光吸收光谱。
裸体和 PC AuNP 不存在和存在时睾酮的体外代谢
进行了初步研究以确定孵育时间和微粒体蛋白质浓度,以提供 TST 的线性代谢率(最终浓度为 10 μM)。 TST 代谢物的产生是线性的,从 1.3 到 9.3 mg 微粒体蛋白 mL -1 最多 60 分钟。代谢测定按照完整描述进行 [25]。简而言之,将微粒体孵育缓冲液中的 pHLM 用 10 μM TST 处理,随后以 0、7、32、63、143、250、400 和 571 μg mL 加入 40 和 80 nm 裸露(无 PC)AuNP −1 ;对于 PC AuNP pHLM 0、7、32、63 和 143 μg mL −1 .微粒体孵育缓冲液含有 100 mM 磷酸盐缓冲液、3.3 mM MgCl 2 和 1 mM EDTA (pH 7.4)。使用和不使用包含 0.25 mM NADP、2.5 mM 6-磷酸葡萄糖和 2 UmL -1 的 NADPH 再生系统启动代谢反应 6-磷酸葡萄糖脱氢酶。在 37°C 下孵育 45 分钟后,加入 4% (v /v ) 磷酸水溶液 (1:1, v /v )。在 3500 rpm 离心 20 分钟后,收集样品上清液并储存在 - 20°C 直至进一步使用。此外,微粒体孵育缓冲液中的单供体 HLM 用 10 μM TST 处理,然后与 63 μg mL -1 孵育 在 37°C 下处理所有裸露和 PC AuNP 45 分钟。孵育结束时,如上处理样品并在 - 20°C 下储存。
标准和样品制备
TST、其代谢物和 13 的一级标准储备液 C3 标记的 TST 作为内标 (ISTD) 在甲醇中制备,浓度为 1 mM,并储存在 - 20°C 直至进一步使用。 TST 及其代谢物的工作标准溶液的浓度为 0.01、0.05、0.1、0.5、1、5、10、50、100 和 200 μM,并连续稀释初级储备溶液。对于标准校准品,将每个工作标准溶液的 50 μL 等分试样添加到 450 μL 反应缓冲液中,以 1:10 稀释,同时还使用 4% 磷酸水溶液制备 0.1 μM ISTD 溶液。质量控制 (QC) 样品的制备浓度分别为 0.01、0.05 和 0.1 μM。
解冻后,样品在室温下以 3500 rpm 离心 20 分钟。上清液中加入 50 μL 的 0.1 μM ISTD,并在 Waters 正压 96 处理器中以 80 psi 的压力处理 Oasis PRIME HLB 96 孔 μelution 板和收集板 1-2 分钟。 (沃特世公司,马萨诸塞州米尔福德)。用 300 μL 含 5% 甲醇的水洗涤并用 50 μL 乙腈/甲醇混合物 (90/10, v /v ),将得到的洗脱液稀释在 50 μL 水中(最终体积为 100 μL)并进行液相色谱-质谱联用 (LC-MS/MS)。
液相色谱-质谱
所有样品均在 Waters UPLC HSS T3 色谱柱(2.1 × 50 毫米,1.8 微米)上进行分离,该色谱柱采用具有三重四极杆检测器 (UPLC TQD) 的 Waters Acquity 超高效液相色谱系统(Waters Corp., Milford, MA)。流动相 A 和 B 分别为 0.1% 甲酸水溶液和 0.1% 甲酸甲醇溶液。使用梯度 LC 方法,流速为 600 μL min -1 0-8.4 分钟。梯度为 0–1 分钟(30% B)、1–3 分钟(至 50% B)、3–3.5 分钟(50% B)、3.5–7 分钟(至 80% B)、7–7.01 分钟(到 98% B)、7.01–7.5 分钟 (98% B) 和 7.51–8.4 分钟 (30% B)。 MS 条件简要描述如下。电离源在电喷雾正离子 (ESI + ) 毛细管电压 4000 V 模式;源温度 150 °C;脱溶剂温度为 450 °C。脱溶剂气 (N2)、锥孔气 (N2) 和碰撞气 (氩气) 的流速为 900 L h -1 , 100 L h −1 , 和 0.1 mL min −1 , 分别。扫描类型为多反应监测 (MRM),MS 运行时间为 8.4 分钟。用于分析的 MRM 离子对在附加文件 1:表 S2 和图 2 中进行了总结。进样量为 2 μL,在整个分析过程中色谱柱保持在 50°C。所有定量方法均基于 0.001 至 20 μM 浓度范围内的七点校准曲线。对 TST 和靶向代谢物的检测限 (LOD) 和定量限 (LOQ) 分别为 0.001 μM 和 0.005 μM。
统计分析
使用 Student's t 评估分散剂对裸露和 PC 对 AuNP 的 DH 和 PDI 的影响 用双尾分布检验。 AuNP 对 pHLM 中 CYP 依赖性 TST 代谢物产生的半数最大抑制浓度 (IC50) 和半数最大活化浓度 (EC50) 是通过使用 GraphPad Prism® 将具有可变斜率的 Hill 方程拟合到观测数据来确定的。使用 GraphPad Prism® 进行单向方差分析 (ANOVA) 以评估 AuNP 处理对单供体 HLM 中 TST 代谢的影响。当效果显着时,在 5% 的显着性水平下使用 Tukey 诚实显着差异 (HSD) 检验进行多重比较。皮尔逊相关系数 (r ) 使用 GraphPad Prism® 6.07 版 (La Jolla, CA) 测定单供体 HLM 的 CYP 活性与 CYP 依赖性 TST 代谢物的产生之间的关系。
结果与讨论
裸体和人血浆蛋白电晕 AuNP 的理化表征
人血浆蛋白电晕 (PC) 对 NP 大小、表面电荷和形态以及光谱特性的影响已使用 DLS、TEM 和 UV-Vis 光谱进行表征(图 1)。 TEM 图像表明,除了 40 和 80 nm PC BPEI-AuNP 外,所有裸露(无 PC)和 PC AuNP 都是单分散的,具有稳定的尺寸分布和独特的 UV-Vis 光谱范围(520-557 nm)(图 1a、b) . TEM 还发现了 PBS 中 AuNP 周围的不同 PC。 40 和 80 nm PC 涂覆的支化聚乙烯亚胺 (BPEI)-AuNP 在 25°C 下 0 分钟在 PBS 中的聚集与尺寸分布的多个峰和吸收光谱相对于裸 BPEI-AuNP 的红移相关联(图 1b)。 25°C 下 0 分钟溶解在 PBS 中以及 37°C 下 0 分钟和 45 分钟溶解在微粒体孵育缓冲液中的 40 和 80 nm PC BPEI-AuNP 的流体动力学直径 (DH) 值不是由 DLS 确定的,以及多个尺寸分布的峰值。 40 nm 裸 BPEI-和 LA-AuNP 和 PC PEG-AuNP 以及 80 nm 裸 BPEI-AuNP 在微粒体孵育缓冲液中的 DH 值在 37°C 下显着增加至 45 分钟,而其值在 80 nm 处降低PC LA-AuNP(表 1)。 40 nm 裸露和 PC PEG-AuNP 以及 80 nm PC PEG-AuNP 的多分散指数 (PDI) 在 37°C 下在 45 分钟时增加。此外,40 和 80 nm 裸 BPEI-AuNP 和 40 nm 裸 PEG-AuNP 的 zeta (z) 电位值随时间显着降低。先前的研究报告称,与人肝微粒体蛋白相关的 AuNP(7 和 70 nm)改变了紫外-可见光范围内的特征吸光度最大值 [21]。这些结果得到了我们实验室最近的研究的支持,即 PC 和人血清白蛋白电晕改变了纳米颗粒的大小、最大吸光度的红移和形态,而与溶解介质和孵育时间无关 [6, 7, 10, 26]。基于 PC 介导的 NP 理化性质和 CYP 活性的酶功能的变化 [6, 7],研究了 40 和 80 nm AuNP 对 CYP 介导的人肝微粒体 TST 代谢的潜在影响相关PC。
<图片>
(a 的透射电子显微照片 ) 去离子水中的 40 和 80 nm 裸 AuNP 和 b 25°C 下 0 小时 PBS 中的 40 和 80 nm PC AuNP、UV 吸收光谱(上部插图)和动态光散射分布(下部插图)。箭头表示 PC 形成。 BPEI 支化聚乙烯亚胺,LA 硫辛酸,PEG 聚乙二醇,ND 未确定,PC 人血浆蛋白电晕,裸体 没有电脑
AuNP 介导的人类肝脏微粒体中的睾酮代谢
总共筛选了 11 种 TST 代谢物,并在 10 μM TST 的混合人肝微粒体 (pHLM) 中发现了 6 种代谢物。图 2 展示了 TST 和六种选定代谢物的靶向 LC-MS/MS 分析。选定的代谢物列表包括五种羟基化 TST 代谢物(2β-OH TST、6β-OH TST、15β-OH TST、16α -OH TST、16β-OH TST)和脱烷基代谢物(雄烯二酮,AD)。这与之前使用人类肝细胞和 HLM 的研究相关,即 TST 主要被羟基化为 6β-OH TST,并在较小程度上羟基化为 2β-OH TST、15β-OH TST、16α-OH TST 和 16β-OH TST 以及脱烷基代谢物,AD [17, 19, 27].
<图片>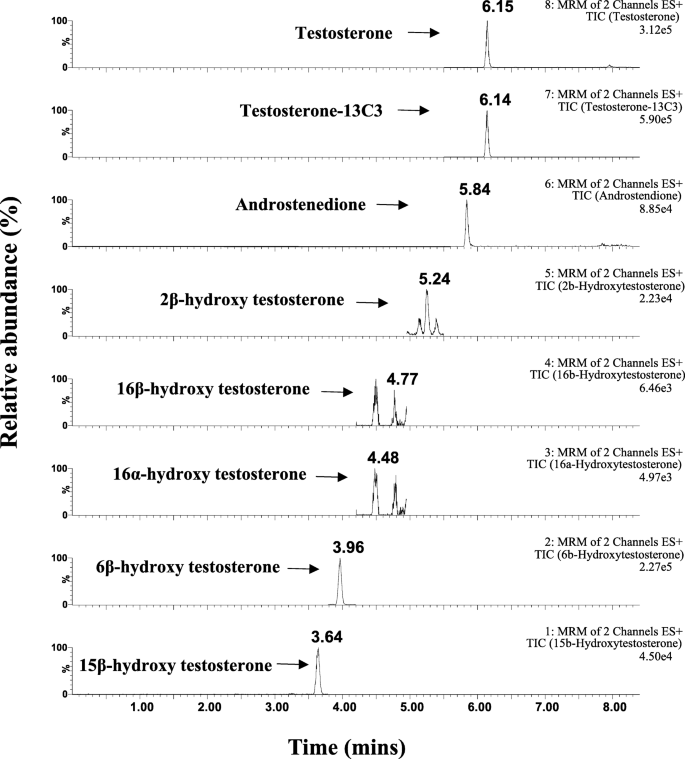
睾酮 (TST) 的提取离子色谱图 (XIC), 13 C3-标记的 TST、雄烯二酮、2β-羟基睾酮 (2β-OH TST)、16α-羟基睾酮 (16α-OH TST)、16β-羟基睾酮 (16β-OH TST)、6β-羟基睾酮 (6β-OH TST) 和 15β -羟基睾酮(15β-OH TST)在混合 HLM 中以 10 μM 睾酮的终浓度在 NADPH 存在下在 37°C 下产生 45 分钟。 高级管理师 人肝微粒体,NADPH 还原型烟酰胺腺嘌呤二核苷酸磷酸
TST 与 pHLM 中的 40 和 80 nm 裸金纳米粒子共孵育的结果如图 1 和图 2 所示。 3 和 4。所有 40 和 80 nm 裸金纳米粒子改变了 pHLM 中 2β-OH TST、6β-OH TST 和 15β-OH TST 的产生,并具有不同程度的抑制(图 3a-f)。 40 nm BPEI-AuNP 产生 6β-OH TST 的半数最大抑制浓度 (IC50) 值为 416 μg mL -1 ;对于 80 nm BPEI-AuNP 438 µg mL −1 ;对于 80 nm PEG-AuNP 387 μg mL -1 (图 3c、d)。对于 15β-OH TST 生产,40 nm BPEI-AuNP 的 IC50 值为 1113 μg mL -1 ;对于 80 nm BPEI-AuNP 415 μg mL −1 ;对于 80 nm PEG-AuNP 480 μg mL -1 (图 3e、f)。这些结果得到了对人癌细胞系和肝组织的体外研究的支持,即金属 NP、AgNP 和多孔硅 NP 阻碍了人上皮性结直肠腺癌 Caco2 细胞、肝细胞癌 HepG2 细胞和人肝微粒体中 6β-OH TST 的产生[22, 28]。除了 40 nm BPEI-AuNP 在最高浓度 (517 μg mL - 1 )(图 4a-d)。 40 和 80 nm BPEI-AuNP 在最高浓度下增加了雄烯二酮 (AD) 的产生,相应的代谢率为 4.3 和 2.1 pmol mg 蛋白质 -1 min −1 , 分别与对照 (0.7 pmol mg protein -1 min −1 )(图 4e、f)。但 80 nm LA-AuNP 抑制 AD 产生,IC50 值为 233 μg mL -1 .这些结果表明,裸金纳米粒子以表面涂层和尺寸依赖性方式介导了所选 TST 代谢物的产生。此外,所有 40 和 80 nm PC AuNP 均不抑制 pHLM 中 TST 的六种选定代谢物的产生,浓度高达 143 μg mL -1 ,与表面涂层无关(图 5 和 6)。特别是,PC 在较高浓度(32 μg mL -1 至 143 μg mL −1 )(图 3c、e 和 5c、e)。该结果表明,40 和 80 nm 裸露的 BPEI-、LA-和 PEG-AuNP 以剂量依赖性方式降低了 TST 羟基化(2β-OH TST、6β-OH TST 和 15β-OH TST)(图 7) )。此外,40 和 80 nm 裸 BPEI-AuNP 增加了 AD 产量,但前者降低了 16β-OH TST。体外研究报道,主要由 CYP3A4 介导的 6β-OH TST 产生被单壁碳纳米管 (SWCNT) 以剂量依赖性方式抑制,但牛血清白蛋白电晕可以缓解它 [17, 29]。我们的实验室最近报道了 40 和 80 nm 裸露和 PC BPEI-AuNP 在细胞和转录水平上作为 CYP1A2、2C9 和 3A4 的抑制剂 [6, 7]。体内研究报道,PEG-AuNP(4 和 13 nm)主要在雄性 BALB/c 小鼠的肝脏中积累,并改变了肝脏 Cyp1a1 和 2b 基因的转录水平 [23]。静脉注射雄性 ICR 小鼠注射 PEG-NH2-AuNP 显示 NP 增加了血浆 TST 水平,而没有精子形态和生育能力 [30]。其他金属 NP、二氧化钛 (TiO2) 在 CD1 雄性小鼠的睾丸中积累,并降低了 cyp1b1 和 2e1 的表达 [31]。流行病学研究报告称,马萨诸塞州不孕不育诊所的成年男性血浆 TST 水平较低,而 3,5,6-三氯-2-吡啶醇 (TCP) 的水平较高,因为毒死蜱 (CFS) 是一种已知的内分泌干扰物和CYP 介导的 TST 代谢抑制剂 [32]。先前的研究报告称,已知的内分泌干扰杀虫剂 CFS、CFS oxon、fonofos、甲拌磷、二乙基甲苯甲酰胺 (DEET) 和氯菊酯显着抑制和/或激活羟基化和/或脱烷基化 TST 代谢物,即 2β-OH 的产生人肝脏中的 TST、6β-OH TST、15β-OH TST 和 AD 和 4-羟基 AD [17]。话虽如此,可以合理地假设 AuNP 可能是一种潜在的内分泌干扰物,通过介导抑制和/或激活对 CYP 介导的 TST 代谢的效力。
<图片>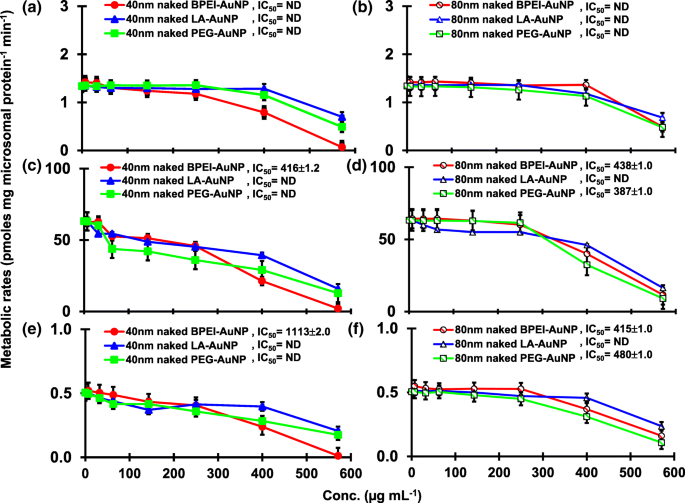
裸(无 PC)AuNP 对 2β-OH TST (a , b ), 6β-OH TST (c , d ) 和 15β-OH TST (e , f ) 在 pHLM 中。数据代表平均值±标准差(n =3)。 集成电路 50,半数抑制浓度; pHLM ,汇集的人肝微粒体; ND , 不是通过使用 GraphPad Prism® 将具有可变斜率的 Hill 方程拟合到观测数据来确定的; 电脑 ,人血浆蛋白; BPEI , 支化聚乙烯亚胺; 洛杉矶 ,硫辛酸; PEG,聚乙二醇; Conc,浓度; 2β-OH TST, 2β -羟基睾酮; 6β-OH TST, 6β -羟基睾酮; 15β-OH TST, 15β -羟基睾酮
<图片>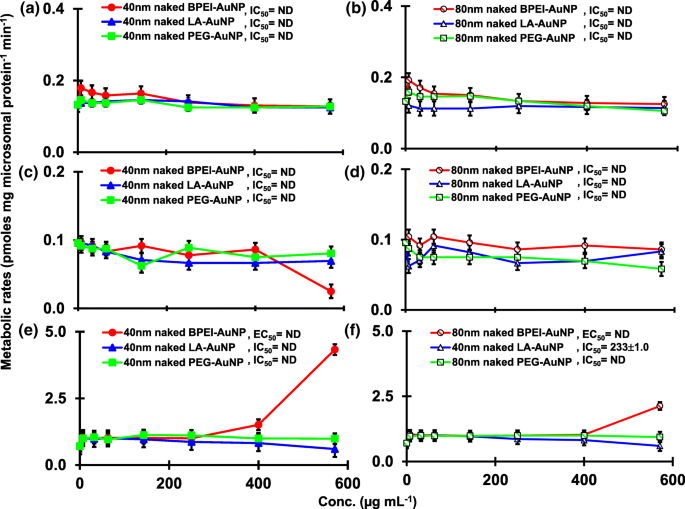
裸(无 PC)AuNP 对 16β-OH TST (a) 产生的抑制和刺激作用 , b ), 16β-OH TST (c , d ) 和 AD (e , f ) 在 pHLM 中。数据代表平均值±标准差(n =3)。 集成电路 50,半数抑制浓度; 欧盟 50,半最大激活浓度; pHLM ,汇集的人肝微粒体; ND , 不是通过使用 GraphPad Prism® 将具有可变斜率的 Hill 方程拟合到观测数据来确定的; 电脑 ,人血浆蛋白; BPEI , 支化聚乙烯亚胺; 洛杉矶 ,硫辛酸; PEG , 聚乙二醇; Conc,浓度; 16α-OH TST, 16α -羟基睾酮; 16β-OH TST, 16β -羟基睾酮; 广告 , 雄烯二酮。
<图片>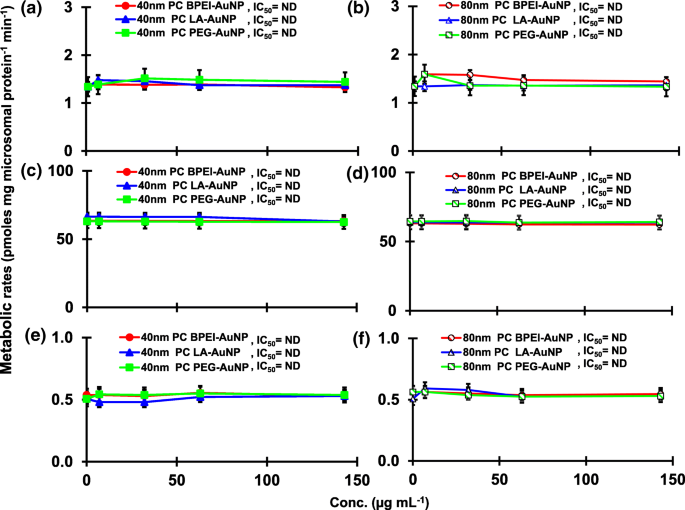
PC AuNP对2β-OH TST(a , b ), 6β-OH TST (c , d ) 和 15β-OH TST (e , f ) 在 pHLM 中。数据代表平均值±标准差(n =3)。 集成电路 50,半数抑制浓度; pHLM ,汇集的人肝微粒体; ND , 不是通过使用 GraphPad Prism® 将具有可变斜率的 Hill 方程拟合到观测数据来确定的; 电脑 ,人血浆蛋白冠; BPEI , 支化聚乙烯亚胺; 洛杉矶 ,硫辛酸; PEG , 聚乙二醇; Conc,浓度; 2β-OH TST, 2β -羟基睾酮; 6β-OH TST, 6β -羟基睾酮; 15β-OH TST, 15β-羟基睾酮。
<图片>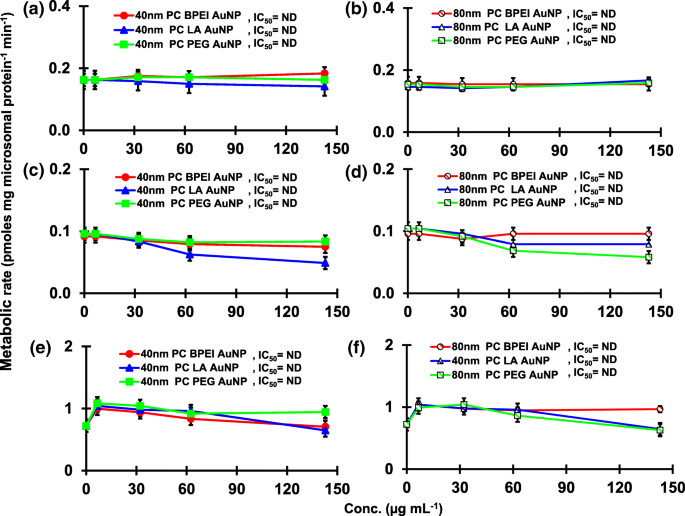
PC AuNP对16α-OH TST(a , b ), 16β-OH TST (c , d ) 和 AD (e , f ) 在 pHLM 中。数据代表平均值±标准差(n =3)。 集成电路 50,半数抑制浓度; pHLM ,汇集的人肝微粒体; ND , 不是通过使用 GraphPad Prism® 将具有可变斜率的 Hill 方程拟合到观测数据来确定的; 电脑 ,人血浆蛋白冠; BPEI , 支化聚乙烯亚胺; 洛杉矶 ,硫辛酸; PEG , 聚乙二醇; Conc,浓度; 16α-OH TST, 16α -羟基睾酮; 16β-OH TST, 16β -羟基睾酮; 广告 , 雄烯二酮。
<图片>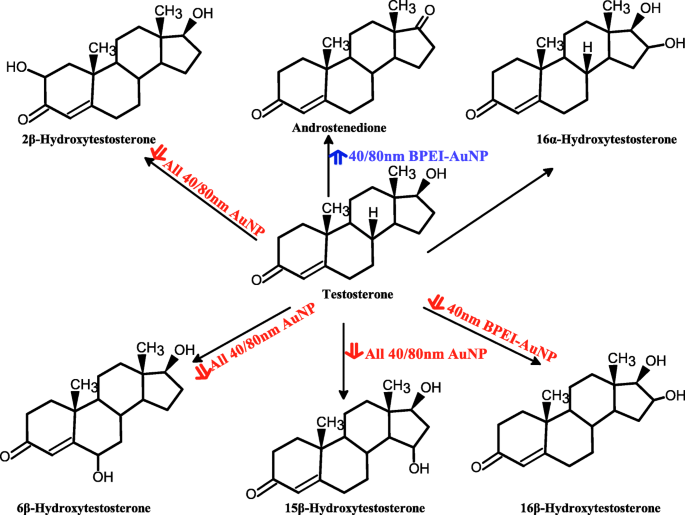
混合的人肝微粒体中睾酮代谢的提议方案和 AuNP 介导的其代谢产物的产生。 AuNP 金纳米粒子,BPEI 支化聚乙烯亚胺,LA 硫辛酸,PEG 聚乙二醇,PC 人血浆蛋白电晕。红色箭头,抑制作用;蓝色箭头,刺激作用
单供体人肝微粒体中的 TST 代谢及其通过 AuNP 的调节
六种选定代谢物的产生特征在于从具有不同程度 CYP 活性的供体中分离出的单一供体 HLM 在基于 pHLM 的非抑制浓度的 40 和 80 nm 裸露和 PC AuNP(10 μg mL -1 )。在三个不同的单一供体 HLM 中观察到 TST 代谢的个体差异(附加文件 1:图 S1)。每种 CYP 酶的催化活性与 TST 衍生代谢物的产生之间的关系在三个不同的单供体 HLM 中表征,这些 HLM 包含低、中和高 CYP 催化活性(附加文件 1:表 S1)。 6β-OH TST的产生与CYP2C19的活性呈正相关(r =0.99 和 p =0.01) 和 CYP3A4 (r =0.99 和 p =0.03) 在个体中(附加文件 1:图 S2)。 AD 产生与 CYP4A11 呈负相关 (r =− 0.98 和 p =0.04)(附加文件 1:图 S3)。这些结果与之前的一项研究一致,即 CYP3A4 和 CYP2D6 分别在主要 TST 代谢物 6β-OH TST 和 AD 的产生中起关键作用 [17]。该研究还表明,TST CYP 依赖性代谢的个体差异取决于 CYP 酶的基因型及其表型活性。先前的一项研究报道,CYP 多态性和表型是 CYP 功能的关键特征,并导致分类药物遗传学表型为弱、中、广泛和超快速代谢者,导致个体对药物不良反应和/或药物疗效和剂量的易感性CYP 酶的弱代谢者,即 CYP3A4 可能对暴露于 CYP 抑制剂 AuNP 的 TST 代谢敏感 [33]。
如图所示。如图 8 和 9 所示,TST 与非抑制性浓度的 AuNP 共培养导致单供体 HLM 中 CYP 介导的 TST 代谢增加和/或减少,这是大小和表面变化修饰的函数。方差分析表明 AuNP 大小的显着变化 (p <0.0001),表面涂层 (p <0.0001) 和 PC 形成 (p <0.0001) 在单供体 HLM(HDA1、HDB2 和 HDC3)中观察到六种选定的 TST 代谢物的产生。
<图片>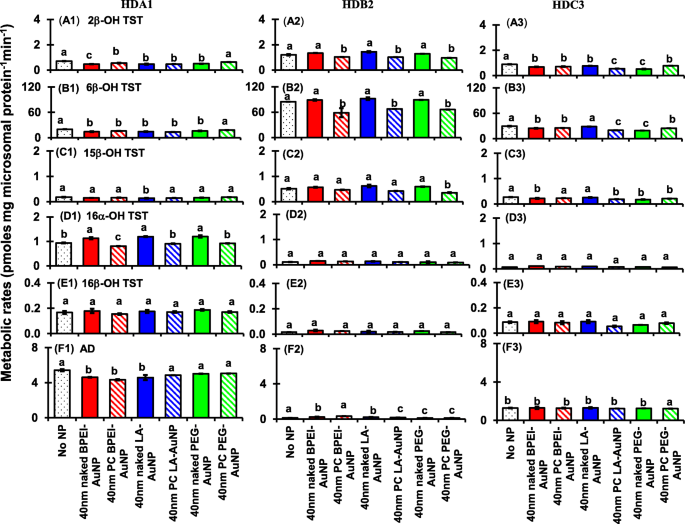
Effects of 40 nm naked and PC AuNP on the production of 2β-OH TST (A1–A3), 6β-OH TST (B1–B3), 15β-OH TST (C1–C3), 16α-OH TST (D1–D3), 16β-OH TST (E1–E3), and AD (F1–F3); in three different single donor HLM (HDA1, HDB2, and HDC3). Means followed by the same letter were not significantly different for each nanoparticle (Tukey’s honest significant difference =5%). Naked no PC, PC human plasma protein corona, HLM human liver microsomes, BPEI branched polyethylenimine, LA lipoic acid, PEG polyethylene glycol, 2β -OH TST 2β-hydroxytestosterone, 6β -OH TST 6β-hydroxytestosterone, 15β -OH TST 15β-hydroxytestosterone, 16α -OH TST 16α-hydroxytestosterone, 16β -OH TST 16β-hydroxytestosterone, AD androstenedione

Effects of 80 nm naked and PC AuNP on the production of 2β-OH TST (A1–A3), 6β-OH TST (B1–B3), 15β-OH TST (C1–C3), 16α-OH TST (D1–D3), 16β-OH TST (E1–E3), and AD (F1–F3); in three different single donor HLM (HDA1, HDB2, and HDC3). Means followed by the same letter were not significantly different for each nanoparticle (Tukey’s honest significant difference =5%). Naked no PC, PC human plasma protein corona, HLM human liver microsomes, BPEI branched polyethylenimine, LA lipoic acid, PEG polyethylene glycol, 2β -OH TST 2β-hydroxytestosterone, 6β -OH TST 6β-hydroxytestosterone, 15β -OH TST 15β-hydroxytestosterone, 16α -OH TST 16α-hydroxytestosterone, 16β -OH TST 16β-hydroxytestosterone, AD androstenedione
All 40 nm naked and PC AuNP decreased both 2β-OH TST and 6β-OH TST production in HDA1 and HDC3 except for PC PEG-AuNP in the former and naked LC-AuNP in the latter, whereas in HDB2, only PC AuNP potentiated inhibition of their productions, irrespective of surface coatings (Fig. 8(A1–B3)). The 40 nm naked LA-AuNP was an inhibitor for 15β-OH TST production in HDA1; for HDB2 PC PEG-AuNP; and for HDC3 naked BPEI- and PEG-AuNP and PC LA- and PC PEG-AuNP (Fig. 8(C1–C3)). All 40 nm naked AuNP were an activator for 16α-OH TST production in HDA1 but PC attenuated it except for PC BPEI-AuNP which potentiated its inhibition (Fig. 8(D1)). All 40 nm naked and PC AuNP did not influence the production of 16β-OH production within individuals (Fig. 8(E1–E3)). AD production was modulated by the 40 nm naked and PC AuNP with varying degrees of inhibition except for PC PEG-AuNP which served an activator in HDC3 (Fig. 8(F1–F3)).
The 80 nm naked and PC AuNP-mediated inhibition for 2β-OH TST production was observed within individuals except for 80 nm naked and PC PEG-AuNP which served as the activators in HDB2 and HDC3, respectively (Fig. 9(A1–A3)). These results were not consistent with all naked and PC 40 nm AuNP-mediated inhibition for 2β-OH TST, irrespective of surface coatings (Fig. 8(A1–A3)). For 6β-OH TST, the 80 nm naked AuNP were the activators except for LA-AuNP but PC potentiated its inhibition in HDB2 (Fig. 9(B2)), which was similar to the 40 nm naked and PC AuNP-mediated inhibition and/or activation, respectively (Fig. 8(B2)). For 15β-OH TST, 80 nm naked BPEI- and PEG-AuNP and PC PEG-AuNP were the activators in HDB2 and in HDC3, respectively (Fig. 9(C2 and C3)). The 80 nm naked AuNP increased 16α-OH TST production in HDA1, irrespective of surface coatings but PC attenuated it except for PC LA-AuNP which was an inhibitor (Fig. 9(D1)). This is similar to the 40 nm naked and PC AuNP-mediated activation and attenuation for 16α-OH TST production in HDA1 (Fig. 8(D1)). All 80 nm naked and PC were not inhibitory to 16β-OH TST production within all individuals, irrespective of surface coatings (Fig. 9(E1–E3)). These results were consistent with the 40 nm naked and PC-mediated its production within individuals (Fig. 8(E1–E3)). For AD production, the 80 nm naked BPEI- and PEG-AuNP were the inhibitors but PC attenuated and vice versa with naked and PC LA-AuNP in HDA1 (Fig. 9(F1)). The 80 nm naked and PC AuNP decreased its production in HDC3 except for PC PEG-AuNP, which was an activator (Fig. 9(F3)). This study strongly suggests that AuNP interaction with CYP enzymes in HLM cause a decrease and/or increase in TST conversion to hydroxylated and dealkylated metabolites within individuals and the presence of PC played the inhibitive or protective role. In vivo study reported that the male CD-1 mice orally administrated with ketoconazole, a noncompetitive CYP3A4 inhibitor showed that a decrease in serum TST level, gonadal TST secretion, and hepatic TST hydroxylation activity that included 6β-OH TST, 15α-OH TST, 15β-OH TST, and 16β-OH TST [34]. In vitro studies with human hepatocyte, C3A cell line, HepG2 cell line, HLM, and recombinant CYP enzymes suggested that AuNP modulated the activity of various CYP enzymes that included CYP1A2, 2C9, 2C19, 2D6, 2E1, and 3A4 [6, 7, 20, 21]. PC and human serum albumin corona mitigated an inhibitory effect of BPEI- and LA-AuNP on CYP1A2, 2C9, and 3A4 enzyme activity in human hepatocytes and C3A cell line [6, 7]. That being said, it may be rational to propose that AuNP interference with CYP enzymes relates individual susceptibility to unexpected toxicological effects that may result in an altered circulating TST level tied to endocrine disrupting substance and/or drug-drug interaction sharing the same CYP enzymes [35].
结论
These studies exhibit that AuNP interaction with PC definitely modulate CYP-dependent metabolism of TST in HLM derived from a large donor pool that better represents the average American population. The 40 nm naked (no PC) AuNP and to a lesser degree 80 nm naked AuNP inhibited TST hydroxylation but activated TST dealkylation at high concentration. Cationic BPEI-AuNP withheld the production of 6β-OH TST and 15β-OH TST in pooled HLM but the presence of a more biologically relevant PC alleviated their adverse effects as function of size and surface charge modification. In most cases, the 40 and 80 nm naked and PC AuNP are essentially inhibitory to TST metabolism in single donor HLM in a surface chemistry-dependent manner at the noninhibitory concentration. In addition, PC PEG-AuNP caused an activation of AD production in HDC3, irrespective of size. These results may indicate that individual variations in AuNP-mediated TST metabolism could be a factor for their toxicity and could be utilized to identify vulnerable subgroup to TST-disrupting NP.
数据和材料的可用性
All data generated or analyzed during this study are included in this article and its supplementary information file.
缩写
- 11β-OH TST:
-
11β-hydroxytestosterone
- 15β-OH TST1:
-
5β-hydroxytestosterone
- 16α-OH TST:
-
16α-hydroxytestosterone
- 16β-OH TST:
-
16β-hydroxytestosterone.
- 2α-OH TST:
-
2α-hydroxytestosterone
- 2β-OH TST:
-
2β-hydroxytestosterone
- 6α-OH TST:
-
6α-hydroxytestosterone
- 6β-OH TST:
-
6β-hydroxytestosterone
- AD:
-
Androstenedione
- AgNP:
-
银纳米粒子
- ANOVA:
-
One-way analysis of variance
- AuNP:
-
金纳米粒子
- BPEI:
-
Branched polyethylenimine
- CFS:
-
Chlorpyrifos
- CYP:
-
Cytochrome P450
- DEET:
-
Diethyltoluamide
- DH :
-
Hydrodynamic diameters
- DI:
-
去离子水
- DLS:
-
动态光散射
- EC50 :
-
Half maximal activation concentration
- EDC/NHS:
-
1-Ethyl-3-(3-dimethylaminopropyl) carbodiimide/N-hydroxysuccinimide
- ESI + :
-
Electrospray positive
- HLM:
-
Human liver microsomes
- HSD:
-
Tukey’s honest significant difference
- IC50 :
-
Half maximal inhibitory concentration
- ISTD:
-
Internal standard
- LA:
-
Lipoic acid
- LC-MS/MS:
-
Liquid chromatography-mass spectrometry
- LOD:
-
Limit of detection
- LOQ:
-
Limit of quantitation
- MRM:
-
Multiple reaction monitoring
- NADP:
-
Nicotinamide adenine dinucleotide phosphate
- NADPH:
-
Reduced NADP
- naked:
-
No PC
- NP:
-
纳米粒子
- PBS:
-
磷酸盐缓冲盐水
- PC:
-
Human plasma protein corona
- PDI:
-
多分散指数
- PEG:
-
聚乙二醇
- pHLM:
-
Pooled human liver microsomes
- QC:
-
Quality control
- SWCNT:
-
Single-walled carbon nanotube
- TEM:
-
透射电子显微镜
- TFF:
-
Tangential flow filtration
- 二氧化钛:
-
二氧化钛
- TST:
-
Testosterone
- UPLC TQD:
-
Ultra performance liquid chromatography system with Triple quadrupole Detector
纳米材料
- 半导体纳米粒子
- 等离子纳米粒子
- 用于化学传感器的金纳米粒子
- 用于改进诊断和治疗应用的多功能金纳米粒子:综述
- 基于局部表面等离子体共振的金纳米生物传感器能够诊断人类布鲁氏菌病,介绍一种快速且经济的方法
- 二氧化钛纳米颗粒对小鼠的潜在肝脏、大脑和胚胎毒性
- 使用聚(4-苯乙烯磺酸-共-马来酸)增强金磁性纳米颗粒的稳定性:用于蛋白质检测的定制光学特性
- 改性超支化聚甘油作为分散剂,用于控制和稳定碳氢化合物中的金纳米粒子
- 用 6-巯基嘌呤和神经元穿透肽修饰的金纳米颗粒促进 SH-SY5Y 细胞生长
- 桔梗皂苷(桔梗)用于金和银纳米颗粒的绿色合成
- 圆形金纳米粒子:粒径和浓度对拟南芥根系生长的影响
- 由金纳米粒子制成的纹身彻底改变了医学诊断