

来自 M2 巨噬细胞的 microRNA-18a 抑制 TGFBR3 以通过 TGF-β 信号通路促进鼻咽癌进展和肿瘤生长
摘要
目标
鼻咽癌(NPC)是一种具有高度转移和侵袭特性的鼻咽疾病。肿瘤相关的替代激活 (M2) 巨噬细胞被证明与 NPC 相关。基于此,本研究旨在探讨M2巨噬细胞microRNA-18a(miR-18a)在NPC中的作用机制及参与作用。
方法
外周血单个核细胞分化为巨噬细胞,巨噬细胞被白细胞介素4极化为M2型。 SUNE-1 和 CNE2 细胞用恢复或耗尽的 miR-18a 或转化生长因子-β III 受体 (TGFBR3) 转染,以探索它们在 TGF-β 信号通路参与的 NPC 进展中的作用。接下来,将SUNE-1和CNE2细胞与经过修复或耗尽的miR-18a或TGFBR3处理的M2巨噬细胞共培养,以了解它们在TGF-β信号通路参与下的NPC中的联合作用。
结果
MiR-18a在NPC细胞中高表达而TGFBR3低表达。 MiR-18a 恢复、TGFBR3 敲低或与 miR-18a 模拟物共培养,或 si-TGFBR3 转染的 M2 巨噬细胞促进 SUNE-1 细胞进展、小鼠肿瘤生长、降低 p-Smad1/t-Smad1 和升高 p- Smad3/t-Smad3。 miR-18a 下调、TGFBR3 过表达或与 miR-18a 抑制剂或 OE-TGFBR3 转染的 M2 巨噬细胞共培养抑制 CNE2 细胞进展、小鼠肿瘤生长、增加 p-Smad1/t-Smad1 并降低 p-Smad3/ t-Smad3.
结论
我们的研究阐明了来自 M2 巨噬细胞的 miR-18a 通过抑制 TGFBR3 以及 Smad1 失活和 Smad3 激活导致裸鼠 NPC 细胞进展和肿瘤生长。
介绍
鼻咽癌(NPC)是一种上皮性恶性肿瘤,倾向于局部浸润和早期远处转移[1]。鼻咽癌患者常主诉第六神经麻痹和霍纳综合征[2]。目前应用的治疗主要是放疗和放化疗联合治疗[3]。不幸的是,放疗和化疗出人意料地伴随着并发症,而对放疗的获得性抵抗阻碍了鼻咽癌的预后[3]。鉴于挖掘潜在靶向治疗的任务是当务之急。
已证明失调的微 RNA (miRNA) 与 NPC 的肿瘤发生、转移、侵袭以及对放疗和化疗的抵抗有关 [1]。作为 miRNA 的一个亚家族,发现 miR-18a 通过抑制生殖器 1 中的形态发生和 mTOR 通路激活来促进 NPC 进展 [4]。除此之外,进一步证实 miR-18a 可通过 DICER1 调节激活 NPC 细胞增殖和转移 [5]。此外,有证据表明 NPC 细胞进展是由 miR-18a 通过 miRNA 生物发生障碍驱动的 [6]。此外,已证明 miR-18a 在 NPC 的转移中起关键作用 [7]。替代激活 (M2) 巨噬细胞是实体和血液恶性肿瘤的重要组成部分,与进展、转移和治疗抵抗有关 [8]。 M2 极化的肿瘤相关巨噬细胞与 NPC 的不良预后有关 [9]。它有趣地记录了 Epstein-Barr 病毒阴性和 Epstein-Barr 病毒阳性 NPC 中 M2 巨噬细胞的差异 [10]。有一项研究概述了 miR-18a 通过诱导 M1 巨噬细胞来抑制结肠癌细胞的肝转移 [11]。转化生长因子-β III 受体 (TGFBR3) 是一种 TGF-β 共受体,可提供 II 型 TGF-β 受体配体以刺激信号传导和细胞表面的平衡,可溶性 TGFBR3 是癌症进展过程中的调节剂 [12]。据报道,低表达的 TGFBR3 可诱导免疫耐受性肿瘤微环境 [13]。相反,TGFBR3 的瞬时过表达诱导人 NPC 细胞凋亡 [14]。据我们所知,miR-181a诱导的M2巨噬细胞极化通过Kruppel样因子6和CCAAT/增强子结合蛋白α轴促进M2巨噬细胞介导的肿瘤细胞转移[15]。
总的来说,尽管许多研究发现了 miR-18a、TGFBR3 和 M2 巨噬细胞在 NPC 中的独立作用,但这三个因素之间的联合相互作用仍然难以捉摸。考虑到开展本研究是为了破译这些因素在NPC中的作用机制和参与。
材料和方法
道德声明
实验经中南大学湘雅三医院伦理委员会批准,符合医学伦理标准。这项研究是在所有捐助者的书面同意下进行的。动物实验符合国家实验动物管理和使用规定的要求。
外周血单核细胞采集
单核细胞通过贴壁法从健康供者的外周血中获得。外周血标本取自中南大学湘雅三医院血液科健康献血者。单核细胞衍生的巨噬细胞是通过外周血单核细胞 (PBMC) 的塑料粘附获得的,该细胞先前通过密度梯度离心 (Ficoll-Paque, GE Healthcare) 从健康供体血液的血沉棕黄层制剂中分离出来。然后,2.0 × 10 6 PBMC 在补充有 10% 人血清(Millipore,Bedford,MA,USA)和青霉素/链霉素的 Dulbecco 改良 Eagle 培养基(DMEM)中在 12 孔板(NalgeNunc,NY,USA)中培养。 PBMCs贴壁2~3 h后,反复去除上清液和悬浮的PBMCs,获得贴壁单核细胞。
巨噬细胞极化
人巨噬细胞集落刺激因子(M-CSF)诱导单核细胞分化为巨噬细胞,白细胞介素(IL)-4诱导单核细胞分化为M2型巨噬细胞。
巨噬细胞的诱导
单核细胞在 20% 胎牛血清 (FBS)-DMEM 中培养,并加入 M-CSF(100 μg,Peprotech,NJ,USA)至终浓度为 100 ng/mL。每 3 天更换一半培养基,然后补充 100 ng/mL M-CSF。培养至第7-8天,收获部分细胞,免疫荧光法[19]检测巨噬细胞表面标志物CD68[16]、CD163[17]、CD206[18],鉴定巨噬细胞。
巨噬细胞的极化
通过将 20 ng/mL IL-4 (Peprotech) 补充到分化培养基中 24 小时,巨噬细胞被极化为 M2 巨噬细胞。一部分 M2 巨噬细胞样品用于流式细胞术检测。样品分为3管:管1是同一批没有IL-4刺激的贴壁巨噬细胞;管2和3是IL-4刺激的贴壁巨噬细胞。上样时将样品稀释至约 10,000 个细胞,并加入非特异性免疫球蛋白 G 以阻断 Fc 受体。然后在样品中加入非特异性同种型抗体、PE 标记的 CD68 抗体和 PE 标记的 CD163 抗体(均来自 Biolegend,CA,USA)。孵育30 min,用0.5%牛血清白蛋白的磷酸盐缓冲液(PBS)冲洗,离心后加入500 μL PBS制成细胞悬液进行检测。
逆转录定量聚合酶链反应
采用逆转录定量聚合酶链反应(RT-qPCR)检测细胞中miR-18a、CCL22、过氧化物酶体增殖物激活受体γ(PPAR-γ)和TGFBR3 mRNA的表达。
通过 Trizol(Invitrogen,Carlsbad,CA,USA)从细胞中提取总 RNA,并通过 Mir-X miRNA First Strand Synthesis Kit(Clontech,Mountain View,CA,USA)逆转录为互补 RNA,用于 miR-18a 和 PrimeScriptTM RT用于 CCL22、PPAR-γ 和 TGFBR3 的 Master Mix Kit(Takara,大连,中国)。 U6 和 3-磷酸甘油醛脱氢酶 (GAPDH) 是 miR-18a、CCL22、PPAR-γ 和 TGFBR3 的上样对照。 SYBR @ 在 PCR 中使用 LightCycler 480 II 系统(Roche Diagnostics,Indiana,USA)中的 Premix Ex Taq™ II(Perfect Real Time)(Takara)。数据计算采用2 -△△CT 方法。 PCR引物见表1。
Western Blot Assay
应用Western blot检测收集细胞中TGFBR3、总(t)-Smad1、磷酸化(p)-Smad1、t-Smad3和p-Smad3蛋白。
提取细胞总蛋白,根据二辛可宁酸试剂盒测定蛋白浓度。在十二烷基硫酸钠聚丙烯酰胺凝胶电泳中将蛋白质样品加载到孔中,并转移到聚偏二氟乙烯 (PVDF) 膜上。 PVDF 膜用脱脂牛奶封闭,并与一抗 TGFBR3(1:2000,R&D Systems,Minneapolis,MN,USA)、t-Smad1(1:1000)、p-Smad1(1:1000,Santa Cruz Biotechnology)一起孵育、t-Smad3 (1:1000)、p-Smad3 (1:1000) 和 GAPDH (1:1000,均来自 Abcam, Cambridge, MA, UK),然后与辣根过氧化物酶标记的二抗一起孵育( 1:500,杰克逊免疫研究实验室,宾夕法尼亚州,美国)。用 Tween 20 用 tris 缓冲盐水洗涤 3 次,膜通过增强的化学发光显色。信号的量化由美国国立卫生研究院 ImageJ 成像完成。信号强度归一化为GAPDH的处理分析软件。
细胞培养和筛选
人鼻咽癌细胞系 CNE2、TW03、C666-1、SUNE-1 和正常人鼻咽细胞系 NP96(中国科学院上海生命科学研究所,中国上海)在 Roswell Park Memorial Institute (RPMI) 培养- 1640 培养基 (Gibco, CA, USA) 含有 FBS (Gibco)、100 μg/mL 青霉素和 100 μg/mL 链霉素,并在 80% 汇合时传代。 RT-qPCR 用于检测 miR-18a 的表达。在这些NPC细胞系中,CNE2和SUNE-1细胞与NP96细胞的miR-18a表达差异最大和最小,因此选择它们进行miR-18a下调或上调分析。
细胞分组和处理
在所有NPC细胞系中,选择在miR-18a表达上与NP96细胞差异最小的SUNE-1细胞。在 Lipofectamine 2000 (Invitrogen) 规范指导下,用 miR-18a 模拟物、miR-18a 模拟物阴性对照 (NC)、si-TGFBR3 或 si-TGFBR3 NC 转染 SUNE-1 细胞。
在所有NPC细胞系中,选择miR-18a表达与NP96细胞差异最大的CNE2细胞,并用Lipofectamine 2000转染miR-18a抑制剂、miR-18a抑制剂NC、过表达(OE)-TGFBR3或OE-TGFBR3 NC (Invitrogen)。
在 Lipofectamine 2000 (Invitrogen) 规范指导下,M2 巨噬细胞转染 miR-18a 模拟物、miR-18a 模拟物 NC、si-TGFBR3、si-TGFBR3 NC、miR-18a 抑制剂、miR-18a 抑制剂 NC、OE-TGFBR3、或 OE-TGFBR3 NC。
M2 巨噬细胞和 NPC 细胞的共培养
采用Transwell小室细胞共培养研究M2巨噬细胞miRNA对NPC细胞的影响。上腔充满了孔径为0.4 μm的M2巨噬细胞,只阻止上腔的细胞通过,而不能阻止细胞分泌的小分子如囊泡、生长因子、营养物等。 下腔用NPC细胞传播。
SUNE-1 和 CNE2 在正常 FBS (Gibco) 中孵育。采用对数生长期细胞进行实验。
SUNE-1 和 CNE2 细胞与 M2 巨噬细胞在 10% FBS-RPMI-1640 培养基(均来自 Gibco)中在 Transwell 插入式细胞培养皿(Coring,Corning,NY,USA)中共培养,孔径为 0.4 μm .
SUNE-1 细胞未与 M2 巨噬细胞共培养,或与 M2 巨噬细胞、miR-18a 模拟转染的 M2 巨噬细胞、miR-18a 模拟 NC 转染的 M2 巨噬细胞、si-TGFBR3 转染的 M2 巨噬细胞或 si 共培养-TGFBR3 NC 转染的 M2 巨噬细胞。
CNE2 细胞未与 M2 巨噬细胞共培养,或与 M2 巨噬细胞、miR-18a 抑制剂转染的 M2 巨噬细胞、miR-18a 抑制剂 NC 转染的 M2 巨噬细胞、OE-TGFBR3 转染的 M2 巨噬细胞或 OE-TGFBR3 共培养NC转染的M2巨噬细胞。
3-(4, 5-Dimethylthiazol-2-yl)-2, 5-Diphenyltetrazolium Bromide 检测
通过 3-(4, 5-二甲基噻唑-2-基)-2, 5-二苯基四唑溴化物 (MTT) 测定法测试细胞活力,这是一种用于测定线粒体脱氢酶活性的比色测定法,可将 MTT 还原为甲臜。
胰蛋白酶消化并以 4 × 10 4 接种到 96 孔板中 细胞/孔,分别在第 0、12、24、36 和 48 小时去除培养基,并补充 MTT 溶液(500 μL,0.5 g/L)。孵育 4 h,弃上清,加入 200 μL 二甲亚砜溶液孵育细胞。在酶标仪(ELX808IU,BioTek,VT,USA)上测量光密度(OD,490 nm)值。每组设6个平行孔。
菌落形成分析
集落形成实验检测鼻咽癌细胞的集落形成能力,反映细胞群体依赖性和细胞克隆增殖能力。
培养24小时,0.25%胰蛋白酶分离,300个细胞接种于35mm培养皿中,每组3个平行孔。每 3 天更换一次培养基,将细胞培养一周并用 5 mL 4% 多聚甲醛固定。之后,将细胞用结晶紫染色液染色并风干。将培养皿倒置,上面叠加一层透明的网格薄膜,在显微镜(Olympus,Tokyo,Japan)下计数菌落数(50个以上)。
划痕测试
通过划痕测试测试细胞迁移。细胞经胰酶消化后接种于每组3个平行孔的6孔板中,培养至90%汇合。然后,将细胞在含有 2% FBS 的培养基中培养,并用 100 μL 尖端划出垂直划痕。倒置显微镜下于第0和24小时对细胞拍照以测量细胞迁移距离。
Transwell 检测
通过Transwell测定测试细胞侵袭和迁移。 Transwell小室的上室预浸入并加入100 μL Matrigel (Coring),用无血清RPMI 1640培养基按1:100稀释。上下室分别加入200 μL和600 μL无血清RPMI 1640培养基。随后,下室加入 600 μL 含 10% FBS 的 RPMI 1640 培养基,而上室加入 200 μL 细胞悬液(12.5 × 10 4 细胞/毫升)。孵育40 h,细胞用结晶紫染色液染色,用棉签擦拭,显微镜下计数通过Matrigel的细胞数。
流式细胞术
流式细胞术检测细胞凋亡和细胞周期分布。
通过碘化丙啶 (PI) 染色评估细胞周期分布。将细胞以 4 × 10 5 接种到 6 孔板中 细胞/孔,并培养至 70-80% 汇合。预冷70%乙醇固定过夜,离心(弃上清),加入RNAase(1 g/L,200 μL)和Triton X-100(2 μL),PI染色液染色30分钟。之后,根据每个阶段(G0/G1阶段、S阶段和G2/M阶段)的不同细胞荧光强度,通过流式细胞仪(BD Bioscience,NJ,USA)在488 nm处检测细胞周期分布。
通过膜联蛋白 V-异硫氰酸荧光素 (FITC) 和 PI 双染色测量细胞凋亡。将细胞重悬于 500 μL 结合缓冲液中,并用 5 μL Annexin V-FITC 染色溶液和 10 μL PI 溶液染色。还通过流式细胞仪 (BD Bioscience) 在 30 分钟内在没有光照的情况下测试细胞凋亡。在散点图上,左下象限 (Q4) 的活细胞为 FITC − /PI − , 右下象限 (Q3) 早期凋亡细胞为 FITC + /PI − , 右上象限(Q2)晚期坏死和凋亡细胞为FITC + /PI + .凋亡率=早期凋亡百分比(Q3)+晚期凋亡百分比(Q2)。
裸鼠肿瘤异种移植
通过建立裸鼠NPC模型观察肿瘤生长。用0.25%胰蛋白酶分离,对数期的SUNE-1和CNE2细胞配置成5×10 7 的单细胞悬液 细胞/毫升。通过显微注射器将细胞悬液(0.2 mL)注射到小鼠右腋下,建立小鼠模型。模型小鼠在特定的无病原体环境中饲养。从第4天开始观察肿瘤生长情况,每4天称重一次。注射后第20天对裸鼠实施安乐死,切除肿瘤,电子天平称重并拍照。
双荧光素酶报告基因检测
采用双荧光素酶报告基因系统确认miR-18a与TGFBR3 mRNA 3'非翻译区(UTR)的结合位点。利用生物预测网站(http://www.microrna.org/microrna/home.do)对miR-18a的靶基因进行分析,发现miR-18a在3′端存在互补结合位点。 TGFBR3 的 UTR。双荧光素酶报告基因检测被用来进一步验证 TGFBR3 是否被 miR-18a 直接靶向。构建了 TGFBR3 3'UTR 结合位点的 pmirGLO-TGFBR3 野生型 (WT) 和 pmirG-LO-TGFBR3 突变型 (MUT)。通过 Lipofectamine 2000 (Invitrogen) 将 TGFBR3-WT 或 TGFBR3-MUT 和 miR-18a 模拟物或模拟物 NC 共转染到 SUNE-1 和 CNE2 细胞中,并孵育 48 小时。使用荧光素酶检测试剂盒(Promega,Madison,WI,USA)进行细胞分析。
统计分析
采用SPSS21.0统计软件(IBM Corp. Armonk, NY, USA)进行分析。数据表示为平均值±标准偏差。通过t分析两组之间的差异 通过单向方差分析 (ANOVA) 检验多个组之间的差异,然后是 Tukey 的事后检验。 P 被认为存在显着差异 <0.05.
结果
M2 巨噬细胞的鉴定
从健康供者的外周血中收集贴壁法富集的单核细胞和M-CSF诱导的单核细胞。 CD68、CD206、CD163免疫荧光检测证实M-CSF体外诱导的PBMCs转化为具有典型分子特征的巨噬细胞,符合我们的要求(图1a)。
<图片>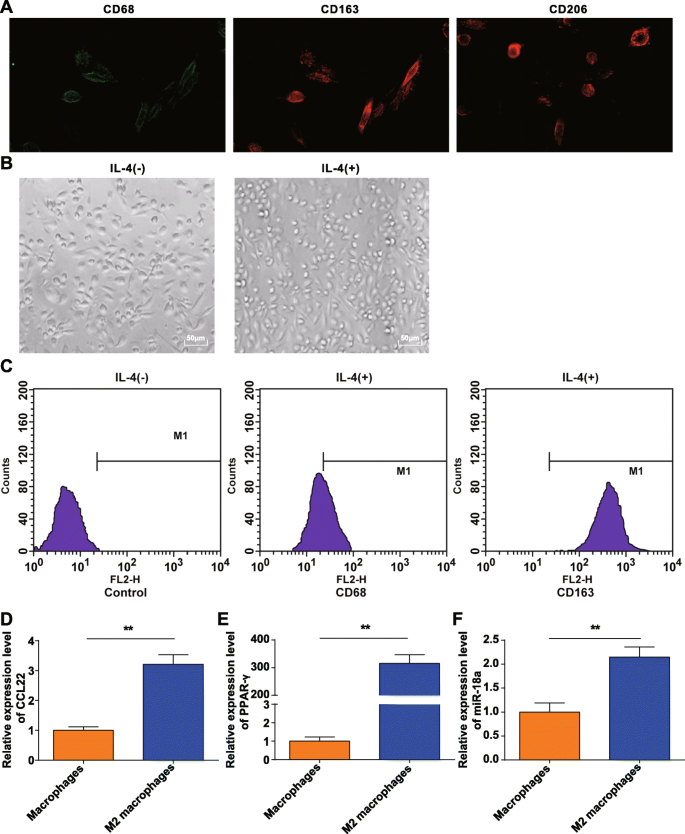
M2巨噬细胞的鉴定。 一 . CD68、CD206 和 CD163 在体外单核细胞诱导后获得的巨噬细胞表面表达。 b IL-4 极化的巨噬细胞是 M2 巨噬细胞。 c CD163 在被 IL-4 极化的 M2 巨噬细胞中高表达而 CD68 低表达。 d CCL22 在 M2 巨噬细胞中高度表达。 e PPAR-γ 在 M2 巨噬细胞中高表达。 f miR-18a 在 M2 巨噬细胞中高表达; *P <0.05; **P <0.01。测量数据表示为平均值±标准差,N =3.通过t分析两组之间的比较 测试
获得的巨噬细胞通过IL-4极化,在显微镜下观察形态。表现为无IL-4刺激的巨噬细胞(M0)多样且不规则,呈圆形、椭圆形或纺锤形。在IL-4的刺激下,M2巨噬细胞变大并主要转向圆形,这符合M2巨噬细胞的形态特征,如前所述[20](图1b)。
流式细胞术检测了 20 ng/mL IL-4 刺激 24 h 的贴壁细胞表面抗原,发现 CD68 的表达率为 21.16%,而 CD163 的表达率为 98.69%(图 1c),证实贴壁细胞是 M2 巨噬细胞。 RT-qPCR显示,相比之下,对于M0细胞,M2巨噬细胞中CCL22和PPAR-γ(典型极化分子)增加(图1d,e),表明M2巨噬细胞成功诱导。
RT-qPCR 还表明,与 M0 巨噬细胞相比,M2 巨噬细胞中的 miR-18a 表达增加(P ˂ 0.05) (图 1f).
在 NPC 细胞中 MiR-18a 高表达而 TGFBR3 低表达
生物信息学网站(miRanda)预测了 miR-18a 的潜在靶标,TGFBR3 被认为是 miR-18a 的一个靶标(图 2a)。实施双荧光素酶报告基因检测以验证 miR-18a 靶向 TGFBR3 的 3'UTR。 TGFBR3-WT 或 TGFBR3-MUT 被克隆到 pmirGLO 载体中,并与 miR-18a 模拟物或 NC 共转染到 SUNE-1 和 CNE2 细胞中。 miR-18a 模拟物对 TGFBR3 3'UTR-MUT 的荧光素酶活性没有影响,但在 SUNE-1 和 CNE2 细胞中削弱了 TGFBR3 3'UTR-WT 的荧光素酶活性,表明 TGFBR3 是受 miR-18a 调控的靶基因(图. 2b).
<图片>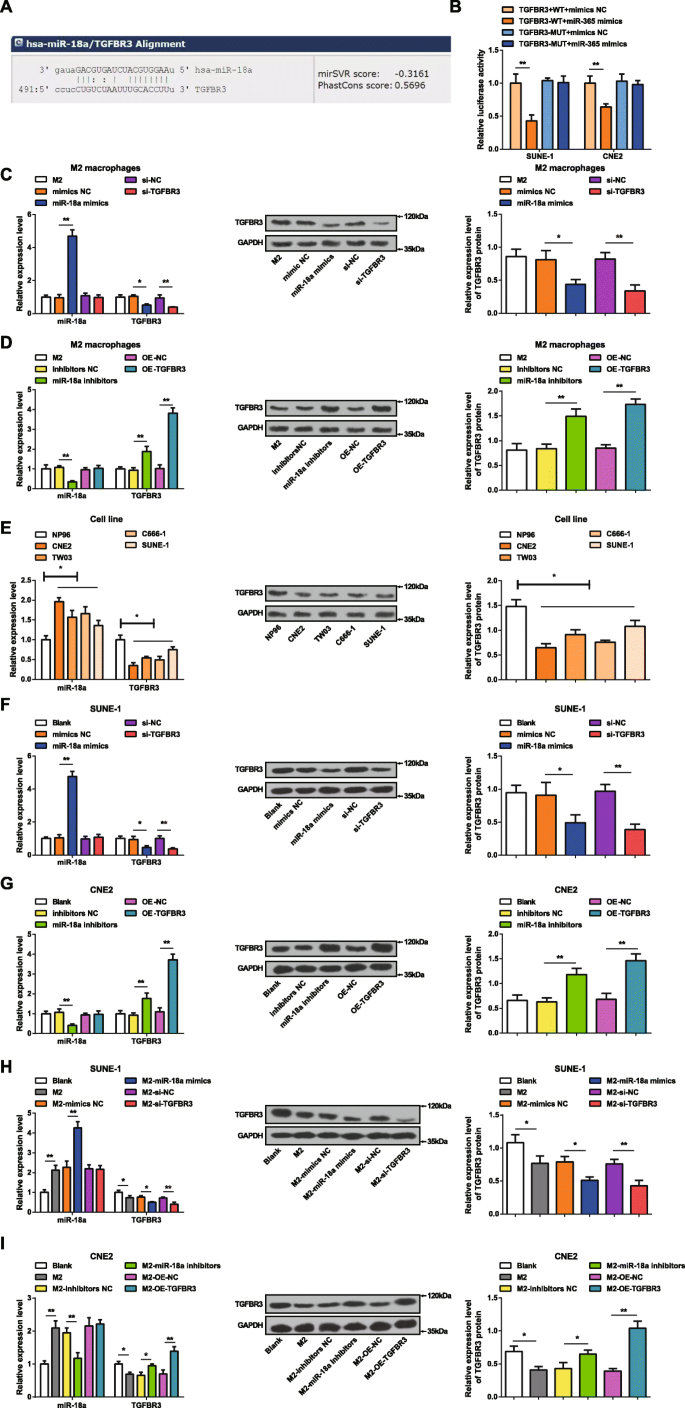
MiR-18a 在 NPC 细胞中高表达而 TGFBR3 表达低。 一 miRanda 预测了靶向 TGFBR3 的 miR-18a。 b 双荧光素酶报告基因检测证实了 miR-18a 靶向 TGFBR3。 c 在 miR-18a 模拟物转染的 M2 巨噬细胞中,miR-18a 表达升高,TGFBR3 表达降低。 d 在转染 miR-18a 抑制剂的 M2 巨噬细胞中,miR-18a 表达降低,TGFBR3 表达升高。 e 与 NP96 细胞相比,NPC 细胞系中 miR-18a 表达升高,TGFBR3 表达降低。 f 在 miR-18a 模拟物转染的 SUNE-1 细胞中,miR-18a 表达升高,TGFBR3 表达降低。 g 在转染 miR-18a 抑制剂的 CNE2 细胞中,miR-18a 表达降低,TGFBR3 表达升高。 h 在与 miR-18a 模拟物转染的 M2 巨噬细胞共培养的 SUNE-1 细胞中,miR-18a 表达升高,TGFBR3 表达降低。 我 在与miR-18a抑制剂转染的M2巨噬细胞共培养的CNE2细胞中,miR-18a表达降低,TGFBR3表达升高; *P <0.05; **P <0.01。测量数据表示为平均值±标准差,N =3.通过t分析两组之间的比较 测试。多组间比较采用单向方差分析,随后采用 Tukey 事后检验
miR-18a 模拟物或 miR-18a 抑制剂在 M2 巨噬细胞中的转染效率通过 RT-qPCR 和蛋白质印迹法测定 M2 巨噬细胞中的 miR-18a 和 TGFBR3 表达来证明。很明显(图2c,d)miR-18a过表达提高了M2巨噬细胞中miR-18a的表达并降低了TGFBR3的表达。相反,抑制 miR-18a 会降低 M2 巨噬细胞中 miR-18a 的表达并增加 TGFBR 的表达。当用 si-TGFBR3 转染 M2 巨噬细胞时,miR-18a 表达未见差异,而 TGFBR3 表达降低。 miR-18a 表达无差异,而 TGFBR3 表达在 OE-TGFBR3 转染至 M2 巨噬细胞后增长。
通过 RT-qPCR 和蛋白质印迹法检测 CNE2、TW03、C666-1、SUNE-1 和 NP96 细胞系中 miR-18a 和 TGFBR3 的表达。在NPC细胞中,与NP96细胞相比,miR-18a表达增加而TGFBR3表达降低(图2e)。由于CNE2细胞和SUNE-1细胞与NP96细胞的miR-18a表达差异最大和最小,因此分别选择它们进行miRNA下调和上调实验。
为了鉴定 miR-18a 和 TGFBR3 对 NPC 细胞的影响,用 miR-18a 模拟物或 si-TGFBR3 转染 SUNE-1 细胞,而用 miR-18a 抑制剂或 OE-TGFBR3 转染 CNE2 细胞。 RT-qPCR 和蛋白质印迹分析表明 miR-18a 模拟 SUNE-1 细胞中升高的 miR-18a 表达和降低的 TGFBR3 表达。 si-TGFBR3 的转染对 miR-18a 的表达没有影响,而在 SUNE-1 细胞中降低了 TGFBR3 的表达。 miR-18a 抑制剂降低了 CNE2 细胞中 miR-18a 的表达并提高了 TGFBR3 的表达。 OE-TGFBR3转染CNE2细胞不影响miR-18a表达,但TGFBR3表达升高(图2f、g)。
为了探索来自 M2 巨噬细胞的 miR-18a 对 NPC 细胞的影响,将 miR-18a 模拟物或 si-TGFBR3 转染的 M2 巨噬细胞或 miR-18a 抑制剂或 OE-TGFBR3 转染的 M2 巨噬细胞与 SUNE-1 共培养或 Transwell 室中的 CNE2 细胞,分别。 RT-qPCR 和蛋白质印迹分析测试了 SUNE-1 或 CNE2 细胞中 miR-18a 和 TGFBR3 的表达。与未转染或转染 miR-18a 模拟物的 M2 巨噬细胞共培养的 SUNE-1 细胞表现出 miR-18a 表达升高和 TGFBR3 表达降低。在与 si-TGFBR3 转染的 M2 巨噬细胞共培养的 SUNE-1 细胞中,miR-18a 表达和 TGFBR3 表达降低没有差异(图 2h)。与未转染的 M2 巨噬细胞共培养后,CNE2 细胞的特点是 miR-18a 表达增加,TGFBR3 表达降低。然而,在与 miR-18a 抑制剂转染的 M2 巨噬细胞预先共培养的 CNE2 细胞中,miR-18a 表达降低,TGFBR3 表达升高。 miR-18a表达无差异,与OE-TGFBR3转染的M2巨噬细胞共培养的CNE2细胞中TGFBR3表达增加(图2i)。
来自 M2 巨噬细胞的 miR-18a 促进 NPC 细胞活力和集落形成能力
应用MTT法和集落形成实验来鉴定miR-18a和TGFBR3对SUNE-1细胞和CNE2细胞活力和集落形成能力的影响。将 miR-18a 模拟物、miR-18a 抑制剂、si-TGFBR3 或 OE-TGFBR3 转染到 SUNE-1 细胞或 CNE2 细胞中。表明在 SUNE-1 细胞中,miR-18a 上调或 TGFBR3 下调增强了细胞活力和增加的集落数(图 3a,e)。在CNE2细胞中,miR-18a抑制或TGFBR3过表达导致细胞活力受损,集落数量减少(图3b、f)。
<图片>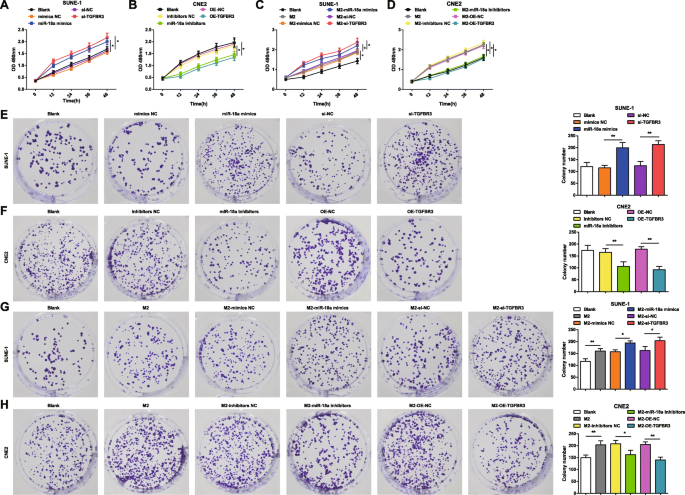
来自 M2 巨噬细胞的 miR-18a 诱导 NPC 细胞活力和集落形成能力。 一 miR-18a 模拟物或 si-TGFBR3 增加了 SUNE-1 细胞的活力。 b miR-18a 抑制剂或 OE-TGFBR3 降低了 CNE2 细胞的活力。 c 与 miR-18a 模拟物或 si-TGFBR3 转染的 M2 巨噬细胞共培养增加了 SUNE-1 细胞的活力。 d 与 miR-18a 抑制剂或 OE-TGFBR3 转染的 M2 巨噬细胞共培养降低了 CNE2 细胞的活力。 e miR-18a 模拟物或 si-TGFBR3 增加了 SUNE-1 细胞的集落数。 f miR-18a 抑制剂或 OE-TGFBR3 减少了 CNE2 细胞的集落数。 g 与 miR-18a 模拟物或 si-TGFBR3 转染的 M2 巨噬细胞共培养增加了 SUNE-1 细胞的集落数。 h 与 miR-18a 抑制剂或 OE-TGFBR3 转染的 M2 巨噬细胞共培养降低了 CNE2 细胞的集落数; *P <0.05; **P <0.01。测量数据表示为平均值±标准差,N =3. 多组间比较采用单向方差分析,随后采用 Tukey 事后检验
为了检查来自 M2 巨噬细胞的 miR-18a 对 NPC 细胞活力和集落形成能力的影响,共培养 miR-18a 模拟物、miR-18a 抑制剂、si-TGFBR3 或 OE-TGFBR3 转染的 M2 巨噬细胞在 Transwell 室中使用 SUNE-1 细胞或 CNE2 细胞。 MTT 和集落形成试验显示,与未转染或 miR-18a 模拟物转染或 si-TGFBR3 转染的 M2 巨噬细胞共培养的 SUNE-1 细胞呈现出增强的细胞活力和增加的集落数量(图 3c,g) .
与未转染的 M2 巨噬细胞共培养的 CNE2 细胞具有增强的细胞活力和增加的集落。然而,与转染miR-18a抑制剂或转染OE-TGFBR的M2巨噬细胞共培养的CNE2细胞表现出细胞活力降低和集落减少(图3d,h)。
来自 M2 巨噬细胞的 miR-18a 促进 NPC 细胞的侵袭和迁移能力
为了更好地理解 miR-18a 和 TGFBR3 如何影响 NPC 细胞的迁移和侵袭,实施了划痕测试和 Transwell 测定。结果表明,转染miR-18a模拟物或si-TGFBR3的SUNE-1细胞的特征是细胞迁移距离和侵袭细胞增加(图4a和5a)。
<图片>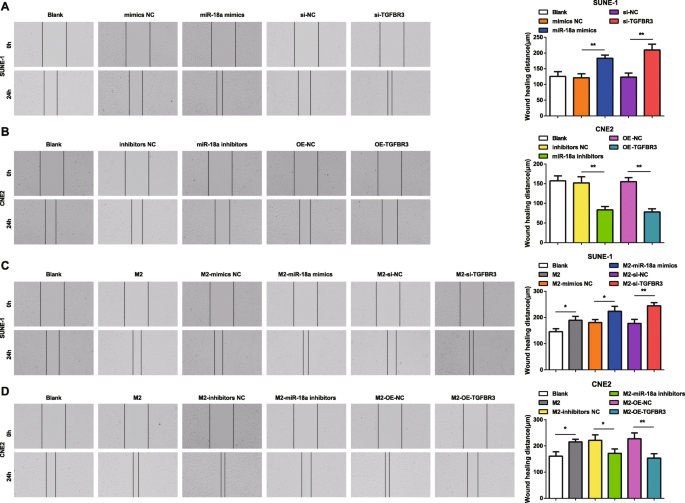
来自 M2 巨噬细胞的 miR-18a 促进 NPC 细胞迁移能力。 一 miR-18a 模拟物或 si-TGFBR3 增加了 SUNE-1 细胞的迁移。 b miR-18a 抑制剂或 OE-TGFBR3 减少了 CNE2 细胞的迁移。 c 与 miR-18a 模拟物或 si-TGFBR3 转染的 M2 巨噬细胞共培养增加了 SUNE-1 细胞的迁移。 d Co-culture with miR-18a inhibitors or OE-TGFBR3-transfected M2 macrophages decreased migration of CNE2 cells; *P <0.05; **P <0.01。 Measurement data were expressed as mean ± standard deviation, N =3. Comparisons among multiple groups were analyzed by one-way ANOVA, followed by Tukey’s post hoc test

miR-18a from M2 macrophages promotes NPC cell invasion ability. 一 miR-18a mimics or si-TGFBR3 increased invasion of SUNE-1 cells. b miR-18a inhibitors or OE-TGFBR3 decreased invasion of CNE2 cells. c Co-culture with miR-18a mimics- or si-TGFBR3-transfected M2 macrophages increased invasion of SUNE-1 cells. b Co-culture with miR-18a inhibitors or OE-TGFBR3-transfected M2 macrophages decreased invasion of CNE2 cells; *P <0.05; **P <0.01。 Measurement data were expressed as mean ± standard deviation, N =3. Comparisons among multiple groups were analyzed by one-way ANOVA, followed by Tukey’s post hoc test
In CNE2 cells transfected with miR-18a inhibitors or OE-TGFBR3, the reductions appeared in cell migration distance and invasion cells (Figs. 4b and 5b).
How miR-18a from M2 macrophages influenced invasion and migration abilities of NPC cells were deciphered by miR-18a mimics-, miR-18a inhibitors-, si-TGFBR3-, or OE-TGFBR3-transfected M2 macrophages co-culturing with SUNE-1 or CNE2 cells. The results highlighted that SUNE-1 cells co-cultured with untransfected, or miR-18a mimics-transfected or si-TGFBR3-transfected M2 macrophages were manifested with increased cell migration distance and invasion cells (Figs. 4c and 5c).
Both cell migration distance and invasion cells increased in CNE2 cells co-cultured with untransfected M2 macrophages. Upon co-culture with miR-18a inhibitors-transfected or OE-TGFBR3-transfected M2 macrophages, CNE2 cells were showed with reduced migration distance and invasion cells (Figs. 4d and 5d).
miR-18a from M2 Macrophages Arrests Fewer NPC Cell at G0/G1 Phase and Suppresses Apoptosis
Cell cycle distribution and apoptosis were tested by flow cytometry to stratify the effects of miR-18a and TGFBR3 on NPC cells. It was indicated that transfection of miR-18a mimics or si-TGFBR3 reduced SUNE-1 cells arrested in the G0/G1 phase, increased cells in the S and G2/M phases, and reduced cell apoptosis rate (Figs. 6a and 7a).

miR-18a from M2 macrophages arrests fewer NPC cell at G0/G1 phase. 一 miR-18a mimics or si-TGFBR3 decreased SUNE-1 cells in the G0/G1 phase, and increased SUNE-1 cells in the S and G2/M phases. b miR-18a inhibitors or OE-TGFBR3 increased CNE2 cells in the G0/G1 phase, and decreased CNE2 cells in the S and G2/M phases. c Co-culture with miR-18a mimics- or si-TGFBR3-transfected M2 macrophages decreased SUNE-1 cells in the G0/G1 phase, and increased SUNE-1 cells in the S and G2/M phases. d Co-culture with miR-18a inhibitors or OE-TGFBR3-transfected M2 macrophages increased CNE2 cells in the G0/G1 phase, and decreased CNE2 cells in the S and G2/M phases; *P <0.05; **P <0.01。 Measurement data were expressed as mean ± standard deviation, N =3. Comparisons among multiple groups were analyzed by one-way ANOVA, followed by Tukey’s post hoc test

miR-18a from M2 macrophages inhibits NPC cell apoptosis. 一 miR-18a mimics or si-TGFBR3 decreased apoptosis of SUNE-1 cells. b miR-18a inhibitors or OE-TGFBR3 increased apoptosis of CNE2 cells. c Co-culture with miR-18a mimics- or si-TGFBR3-transfected M2 macrophages decreased apoptosis of SUNE-1 cells. d Co-culture with miR-18a inhibitors or OE-TGFBR3-transfected M2 macrophages increased apoptosis of CNE2 cells; *P <0.05; **P <0.01。 Measurement data were expressed as mean ± standard deviation, N =3. Comparisons among multiple groups were analyzed by one-way ANOVA, followed by Tukey’s post hoc test
Upon transfection with miR-18a inhibitors or OE-TGFBR3, CNE2 cells in the G0/G1 phase trended toward an elevation while those in the S and G2/M phases toward a reduction, and cell apoptosis rate raised (Figs. 6b and 7b).
With the purpose to decode the mechanism of miR-18a from M2 macrophages in NPC cell cycle distribution and apoptosis, M2 macrophages transfected with miR-18a mimics, miR-18a inhibitors, si-TGFBR3, or OE-TGFBR3 were co-cultured with SUNE-1 cells or CNE2 cells in the Transwell chamber. Co-cultured with untransfected, or miR-18a mimics-transfected or si-TGFBR3-transfected M2 macrophages, reduced SUNE-1 cells were displayed in the G0/G1 phase and increased cells in the S and G2/M phases, and SUNE-1 cell apoptosis rate decreased (Figs. 6c and 7c).
Co-cultured with untransfected M2 macrophages, CNE2 cells in the G0/G1 phase reduced, cells in the S and G2/M phases increased, and apoptosis rate declined. On the contrary, co-cultured with miR-18a inhibitors-transfected or OE-TGFBR3-transfected M2 macrophages, CNE2 cells in the G0/G1 phase elevated while those in the S and G2/M phases decreased, and apoptosis rate elevated (Figs. 6d and 7d).
miR-18a from M2 Macrophages Reduces p-Smad1/t-Smad1 and Elevates p-Smad3/t-Smad3 in NPC Cells
Western blot assay detected TGF signaling pathway-related proteins in NPC cells to further explain the effects of miR-18a and TGFBR3 on TGF signaling pathway.
It was explained that transfection of miR-18a mimics or si-TGFBR3 reduced p-Smad1/t-Smad1 and elevated p-Smad3/t-Smad3 in SUNE-1 cells (Fig. 8a).

miR-18a from M2 macrophages decreases p-Smad1/t-Smad1 and increases p-Smad3/t-Smad3 in NPC cells. 一 miR-18a mimics or si-TGFBR3 in SUNE-1 cells decreased p-Smad1/t-Smad1 and elevated p-Smad3/t-Smad3. b miR-18a inhibitors or OE-TGFBR3 in CNE2 cells increased p-Smad1/t-Smad1 and decreased p-Smad3/t-Smad3. c Co-culture with miR-18a mimics- or si-TGFBR3-transfected M2 macrophages decreased p-Smad1/t-Smad1 and elevated p-Smad3/t-Smad3. d Co-culture with miR-18a inhibitors or OE-TGFBR3-transfected M2 macrophages increased p-Smad1/t-Smad1 and decreased p-Smad3/t-Smad3; *P <0.05; **P <0.01。 Measurement data were expressed as mean ± standard deviation, N =3. Comparisons among multiple groups were analyzed by one-way ANOVA, followed by Tukey’s post hoc test
Transfection with miR-18a inhibitors or OE-TGFBR3 led to declined p-Smad3/t-Smad3 and increased p-Smad1/t-Smad1 in CNE2 cells (Fig. 8b).
miR-18a from M2 macrophages influencing TGF signaling pathway in NPC cells was determined by Western blot assay through testing TGF signaling pathway-related proteins in SUNE-1 cells and CNE2 cells which had co-cultured with miR-18a mimics-, miR-18a inhibitors-, si-TGFBR3-, or OE-TGFBR3-transfected M2 macrophages in the Transwell chamber.
SUNE-1 cells co-cultured with untransfected, miR-18a mimics-transfected, or si-TGFBR3-transfected M2 macrophages were manifested with reduced p-Smad1/t-Smad1 and incremental p-Smad3/t-Smad3 (Fig. 8c).
Co-cultured with untransfected M2 macrophages, CNE2 cells trended toward declined p-Smad1/t-Smad1 and elevated p-Smad3/t-Smad3. In an opposite way, CNE2 cells were demonstrated with increased p-Smad1/t-Smad1 and decreased p-Smad3/t-Smad3 when co-cultured with miR-18a inhibitors-transfected or OE-TGFBR3-transfected M2 macrophages (Fig. 8d).
miR-18a from M2 Macrophages Induces Tumor Growth in Nude Mice with NPC
Tumor xenografts were conducted on nude mice to further elucidate the impacts of miR-18a and TGFBR3 on tumor growth of NPC.
It was indicated that injected with miR-18a mimics-transfected or si-TGFBR3-transfected SUNE-1 cells, mice were manifested with enlarged tumor volume and heavier tumor weight (Fig. 9a).

miR-18a from M2 macrophages promotes tumor growth in nude mice with NPC. 一 miR-18a mimics or si-TGFBR3 in SUNE-1 cells increased tumor volume and weight. b miR-18a inhibitors or OE-TGFBR3 in CNE2 cells decreased tumor volume and weight. c Co-culture with miR-18a mimics- or si-TGFBR3-transfected M2 macrophages increased tumor volume and weight. d Co-culture with miR-18a inhibitors or OE-TGFBR3-transfected M2 macrophages decreased tumor volume and weight; *P <0.05; **P <0.01。 Measurement data were expressed as mean ± standard deviation, three nude mice in each group. Comparisons among multiple groups were analyzed by one-way ANOVA, followed by Tukey’s post hoc test
Reduced tumor volume and weight were presented in mice with injection of miR-18a inhibitors-transfected or OE-TGFBR3-transfected CNE2 cells (Fig. 9b).
Tumor growth was observed in mice which had injected with miR-18a mimics-, miR-18a inhibitors-, si-TGFBR3-, or OE-TGFBR3-transfected M2 macrophages to illustrate the mechanism of miR-18a from M2 macrophages in NPC.
After co-culture with untransfected, miR-18a mimics-transfected or si-TGFBR3-transfected M2 macrophages, SUNE-1 cells were injected into mice and mice were observed with larger tumor volume and heavier tumor weight (Fig. 9c).
CNE2 cells were co-cultured with M2 macrophages and injected into mice with the results suggesting growing tumor volume and weight. Both tumor volume and weight were inclined to reduce when mice were injected with CNE2 cells which had co-cultured with miR-18a inhibitors-transfected or OE-TGFBR3-transfected M2 macrophages (Fig. 9d).
Discussion
NPC refers to a polygenic disease threatened by a wide range of factors [21]. MiRNAs are previously implied to participate in the pathogenesis of NPC via regulation of their target genes which are indicators of cellular processes and pathways [22]. Concretely, miR-18a advances NPC progression by miRNA biogenesis impairing [6]. Given that this study goes forward to decipher the combined interactions of miR-18a from M2 macrophages and TGFBR3 in NPC with the conclusion elucidating that miR-18a from M2 macrophages stimulates NPC progression via TGFBR3 inhibition (Fig. 10).

Schematic representation of macrophage-derived exosomal miR-18a in NPC and the involvement of TGFBR3-mediated TGF-β signaling pathway
At the start of this study, macrophages are stimulated by IL-4 to differentiate to M2 macrophages which are found to enrich the expression of miR-18a. As we know, during macrophage polarization, miRNA’s expression was altered [23]. In addition, M2 polarization enriches genes which are involved in the cell cycle and metabolic processes, and the M2 phenotype is conducive to tumor growth and angiogenesis in neoplastic tissues [24]. Based on the M2 macrophages-enriched miR-18a, a series of experiments were successfully conducted.
Initially, our study has uncovered that miR-18a is highly expressed while TGFBR3 is poorly expressed in NPC cells. Drawn from a previous study, it is concluded that miR-18a is overexpressed in NPC tissues with its association with lymph node metastasis and clinical stage [5]. Besides that, miR-17-92 cluster members including miR-18a are documented to be overexpressed in NPC tissues [25]. Furthermore, upregulated miR-18a is reported to demonstrate in NPC tissues which is connected with tumor node metastasis stage and tumor size [4]. Experimentally, except for the downregulated TGFBR3 in tongue squamous cell carcinoma [26], there has been another study depicting reduced TGFBR3 in clear-cell renal cell carcinomas accompanied by unwanted prognosis [27]. Anyhow, the results in this study are consistent with these study findings to some extent.
In order to explore the roles of miR-18a and TGFBR3 in NPC cell progression, we have conducted a series of assays with the results indicating that upregulated miR-18a or downregulated TGFBR3 triggers NPC cell progression while miR-18a repression or TGFBR3 elevation has the opposite effects on NPC cells. Widely, suppression of miR-18a is evidenced to hamper cell progression in malignancies including ovarian cancer, colitis-associated colorectal cancer, and hepatocellular carcinoma [28,29,30]. Narrowly, an existed study has pronounced that upregulated miR-18a promotes NPC cell progression via mediation of DICER1 [6]. In addition, it is noticed that overexpressed miR-18a in NPC is believed to connect with NPC metastasis and repressed miR-18a partially contributes to better prognosis of NPC patients [31]. Lately, it is surveyed that downregulation of miR-18a is capable of discouraging NPC proliferation, invasion, and migration [4]. Additionally, a decrease in TGFBR3 expression is regarded to link with laryngeal squamous cell carcinoma (LSCC) invasion and miR-223/TGFBR3 axis regulation takes part in LSCC progression inhibition [32]. TGFBR3 elevation is documented to restrict NPC cell viability, induce apoptosis, and activate pro-apoptosis signaling pathways [14]. Previously, a study has indicated that upregulation of TGFBR3 promotes apoptosis and cells arrested in the G2/M phase, resulting in impaired cell viability and migration in salivary gland adenoid cystic carcinoma [33]. Intriguingly, it is formerly described that induction of TGFBR3 contributes to disrupt intrahepatic cholangiocarcinoma progression [34].
Despite the protective role of lowly expressed miR-18a and overexpressed TGFBR3 in NPC cell in vitro, we have performed tumor xenografts in nude mince in vivo for further verification with the results explaining that miR-18a knockdown or TGFBR3 elevation restrains tumor growth in nude mice. As demonstrated in a prior study, miR-18a-injected nude mice show with enhanced tumor growth [5] and conversely, the miR-18a antagomir-injected nude mice are displayed with suppressed tumor growth in NPC [4]. In the light of the TGFBR3 reduction in tumor growth, it is suggested that poorly expressed TGFBR3 provokes tumor formation in clear-cell renal cell carcinoma [27]. In the opposite way, an increase in TGFBR3 is recognized to hinder tumor growth in lung cancer with the presence of long non-coding RNA ADAM metallopeptidase with thrombospondin type 1 motif, 9 antisense RNA 2 elevation, and miR-223-3p suppression [35]. This study has also predicted and verified that TGFBR3 is a target gene of miR-18a. But, more studies still need to be conducted for further verification.
Conclusion
Generally speaking, this study elaborates the concrete mechanisms that miR-18a from M2 macrophages inhibits TGFBR3 expression to exacerbate the progression of NPC via TGF-β signaling pathway, the results of which is abrogated by miR-18a knockdown or TGFBR3 elevation. This study updates the therapeutic target for NPC. However, a large cohort researches are still in need for in-depth explorations.
数据和材料的可用性
不适用。
纳米材料
- 通过溶胶-凝胶工艺制备的纳米结构二氧化硅/金-纤维素-键合氨基-POSS 混合复合材料
- 通过聚乙二醇化脂质体改善蟾蜍灵的抗肿瘤功效和药代动力学
- 钯(II)离子印迹聚合物纳米球的制备及其从水溶液中去除钯(II)
- 5-氨基乙酰丙酸-角鲨烯纳米组件用于肿瘤光检测和治疗:体外研究
- 用 6-巯基嘌呤和神经元穿透肽修饰的金纳米颗粒促进 SH-SY5Y 细胞生长
- 磁赤铁矿纳米颗粒作为纳米酶,改善甘蓝型油菜的生长和非生物胁迫耐受性
- 改性 BiOCl 的合成和表征及其在从水溶液中吸附低浓度染料中的应用
- 利用柠檬汁通过水热反应制备的荧光碳量子点的材料和光学特性
- InGaAs/InP 核壳纳米线的自种 MOCVD 生长和显着增强的光致发光
- CpG 的改性 Fe3O4 磁性纳米颗粒递送抑制肿瘤生长和自发性肺转移以增强免疫治疗
- 纳米藻酸盐通过反胶束合成:阿霉素封装和乳腺癌细胞毒性
- 空心聚苯胺微/纳米球的制备及其从废水中去除 Cr (VI) 的能力