

用于细胞内蛋白质递送的二氧化硅纳米颗粒:一种使用绿色荧光蛋白的新型合成方法
摘要
在这项研究中,提出了一种制备具有窄尺寸分布的绿色荧光蛋白 (GFP) 掺杂二氧化硅纳米粒子的新方法。由于其自发荧光,GFP 被选为模型蛋白。蛋白质掺杂的纳米粒子在细胞内蛋白质递送领域具有很高的应用潜力。此外,荧光标记的颗粒可用于生物成像。使用多步合成过程将这些蛋白质掺杂的纳米粒子的大小从 15 纳米调整到 35 纳米,包括粒子核合成和壳再生步骤。 GFP 通过一锅反应选择性地掺入核或壳或两者的二氧化硅基质中。通过测定粒径、流体动力学直径、ζ-电位、荧光和量子产率来表征获得的纳米颗粒。测量结果表明,GFP 的荧光在粒子合成过程中得以保持。细胞摄取实验表明,GFP 掺杂的纳米粒子可用作稳定有效的荧光探针。该研究揭示了所选方法将功能性生物大分子掺入二氧化硅纳米粒子的潜力,从而开辟了细胞内蛋白质递送等新应用领域。
背景
近年来,由于诸如生物传感器 [1] 或生物反应器 [2] 等材料的广泛应用潜力,以及在受控蛋白质递送领域 [3],将蛋白质封装到微米和纳米颗粒中得到了广泛的关注。细胞内蛋白质递送 [4] 和组织工程 [5]。在许多这些应用中,封装酶的催化活性是此类材料的一项基本功能。相比之下,药物相关蛋白质、肽激素或抗体作为此类纳米材料的潜在货物,通过与组织或细胞内的靶标进行特异性结合来发挥其功能。因此,所有这些应用的一个先决条件是保持货物蛋白的完整构象和功能。纳米结构系统由于其体积小、比表面积大和其他独特的特性,已成为生物医学研究中发展最快的领域之一[6]。因此,开发新的颗粒载体以提高设计系统的功能和稳定性是该领域的一个重要课题 [7]。纳米颗粒载体的基质可以基于生物大分子或有机成分,如碳水化合物、脂质或聚合物,形成固体脂质纳米颗粒、脂质体或树枝状聚合物等系统。此外,纳米结构系统也可以基于无机材料,如金属或氧化物 [8]。所有这些材料系统都必须满足各种常见和特定的要求。首先,基质材料必须具有生物相容性以促进安全应用[9]。其次,它们必须足够稳定,才能在系统的整个生命周期中发挥其作为载体材料的作用。此外,它们必须提供显着的蛋白质负载和保留能力以及控制蛋白质释放的能力[10]。
除了通过吸附或共价结合将蛋白质附着到纳米物体的表面 [11] 之外,蛋白质还可以被包裹在纳米结构中,从而提高它们的稳定性和酶活性 [2]。纳米捕获可以通过溶胶-凝胶处理[12] 或油包水微乳液方法通过二氧化硅前体的水解和缩合来实现,导致在水-油界面处围绕壳的酶聚合[13]。在这些方法中,蛋白质的截留可以通过两种不同的化学方法发生,使用共价或非共价结合过程 [14]。特别是,无定形二氧化硅因其高生物相容性、惰性和机械稳定性而成为一种很有前途的蛋白质载体材料 [15]。已经遵循了各种途径,尤其是将酶包封到二氧化硅中的仿生方法 [2, 16],其中酶的释放曲线由接头的化学反应或二氧化硅基质的降解控制。介孔材料也被用作基质,将酶固定在 2-50 nm 的孔内 [13, 17]。可以通过使用“看门人”策略或修改孔的内表面来控制与药物的结合亲和力来调整介孔纳米粒子的货物释放 [10b]。然而,孔径可能会限制酶加载到所执行的介孔二氧化硅支架中 [18],这就是为什么最近正在研究蛋白质递送的新策略的原因。
由于二氧化硅纳米粒子广泛用于生物成像 [19],荧光蛋白的掺入构成了生成生物相容性荧光探针的一种选择。例如,文献 [20] 中描述了通过反乳化技术将绿色荧光蛋白 (GFP) 掺入二氧化硅纳米粒子中。这些研究表明,将 GFP 掺入二氧化硅颗粒基质中不仅可以增强蛋白质的荧光强度,还可以增强其热稳定性、化学变性和蛋白酶处理的稳定性。然而,该方法不太适合合成具有窄尺寸分布的较低纳米级范围内定义明确的二氧化硅纳米粒子。此外,合成条件包括接触表面活性剂、醇或强碱以及高温,这些都可能与易感蛋白质的掺入不相容[20, 21]。
因此,我们报告了一种使用 GFP 作为模型蛋白制备掺杂蛋白质的二氧化硅纳米粒子的新方法。为此,我们在温和的合成条件(室温、低盐度)下使用一锅法合成,然后进行透析纯化。该方法的特点是有可能制备在低于 50 nm 的尺寸范围内表现出窄尺寸分布的蛋白质包埋二氧化硅纳米颗粒。
方法
材料
所有化学品均购自 Sigma-Aldrich (Taufkirchen, Germany),未经进一步纯化。所有合成和纯化步骤均使用超纯水(18.2 MΩ,Milli-Q 水纯化系统 ELIX 20 型,美国密理博公司)。
GFP的准备
如别处所述,通过蛋白质表达和随后的纯化获得 GFP [22]。简而言之,使用基于 pQE 载体系统(Qiagen,Hilden,Germany)的高水平细菌表达载体在大肠杆菌中表达包括 N 端 His6 标签的 GFP。大肠杆菌 XL1-Blue 并通过带镍亲和层析(Qiagen,Hilden,Germany)纯化。随后,将蛋白质转移到浓缩器装置(3 kDa 截留分子量(MWCO)膜,Pall,Dreieich,Germany)进行缓冲液交换。通过分别加入 15 mL ʟ-精氨酸和碳酸氢钠溶液将 GFP 洗涤 3 次,随后在 3 mL ʟ-精氨酸/碳酸氢钠溶液中回收。之后,将 GFP 悬浮液通过无菌 0.22 μm 醋酸纤维素过滤器(Carl Roth,Karlsruhe,Germany)过滤到无菌管中。使用前,将蛋白质浓度调整为 1 mg mL -1 在 7.2 mmol L −1 ʟ-精氨酸 (pH =10.3) 或 10.0 mmol L −1 NaHCO3 (pH =9.2) 溶液。
纳米粒子的合成与纯化
根据先前描述的修改方案制备二氧化硅纳米粒子 [23]。简而言之,作为非极性前体的四乙氧基硅烷(TEOS)在双相水/环己烷体系中由ʟ-精氨酸催化水解。
核心粒子的制备
在三颈圆底烧瓶中,将 91 mg (0.52 mmol) ʟ-精氨酸溶解在 69 mL 水中,然后加入 4.5 mL 环己烷作为顶层。在搅拌下将反应混合物加热至 40°C。加入 5.5 mL (24.63 mmol) TEOS 后,将混合物在这些条件下再保持 20 小时。
二氧化硅壳层
对于随后的壳生长步骤,使用核颗粒或由第一壳生长步骤产生的颗粒。对于壳生长,将 14 mg (0.08 mmol) ʟ-精氨酸溶解在 36 mL 水中,并加入 10 mL 先前制备的颗粒分散体。添加 5 mL 环己烷后,将混合物加热至 40°C。加入 3.52 mL (15.8 mmol) TEOS 后,将混合物再搅拌 20 小时。
GFP 掺杂纳米粒子的制备。 为制备 GFP 掺杂的纳米粒子,加入 TEOS 后 30 分钟,加入 200 μg (6.9 nmol) GFP。
粒子净化
通过随后使用纤维素水合物膜(Nadir-透析管,MWCO 10 kDa,Carl Roth,Karlsruhe,Germany)对水(4L,30、90和180分钟后的水交换)透析4小时来纯化纳米颗粒。最后,使用无菌 0.22 µm 醋酸纤维素膜过滤器(Carl Roth,Karlsruhe,Germany)将纳米颗粒过滤到无菌烧瓶中。
透射电子显微镜 (TEM)
使用 JEM-2100F 显微镜(JEOL,Freising,德国)测定形态和平均粒径。使用 X-ImageJ 软件(版本:1.45 s,来自美国国立卫生研究院的 winPenPack X-ImageJ Launcher (http://rsb.info.nih.gov/ij))对 50 个纳米颗粒的随机样本测定粒度分布/).
流体动力学直径
使用 Zetasizer Nano ZSP(Malvern Instruments,Herrenberg,Germany)记录纳米颗粒的流体动力学直径。在测量之前,将粒子分散体按 1:10 稀释在水中。在 25°C 下进行测量。每个样品测量3 × 15次。通过体积分布的计算确定直径。这是使用米氏理论从强度大小分布转换而来的。
ζ-势
使用与上述条件相同的仪器测量 ζ 电位,不同之处在于将样品稀释在 0.01 M KCl (9:1) 中。
分析超速离心 (AUC)
为了测量沉降速度,使用带有 AnTi60 转子的改进型 Beckman-Coulter XL-80 K。对于实验,温度设置为 20 °C,速度设置为 10,000 rpm,并进行了 21 次扫描。二氧化硅的波长设置为 261 nm,GFP 检测的波长设置为 488 nm。
荧光光谱
使用 Fluoromax-3 荧光分光计(Spex,Horiba Scientific,Oberursel,Germany)记录纳米颗粒、纯 GFP 和来自浸出实验的滤液的荧光光谱。对于测量,纯 GFP、颗粒分散体和滤液在水中按 1:10 稀释。激发波长设为488 nm,在498~800 nm光谱范围内记录光谱。
荧光量子产率
使用威廉姆森等人的相关方法确定获得的纳米颗粒和纯 GFP 的量子产率。 [24]。作为 GFP 的参考,使用了罗丹明 6G 和 Atto488。使用与参考染料混合的非掺杂纳米粒子进行比较测量。使用 450 nm 的激发波长记录荧光光谱。使用 Varian Cary 300 Scan UV(Agilent Technologies,Darmstadt,Germany)进行额外的 UV/vis 测量。
对于量子产率的计算,方程。 2 被使用了。
$$ {\varPhi}_P={\varPhi}_S\bullet \frac{{\mathrm{slope}}_S}{{\mathrm{slope}}_P}\bullet {\left(\frac{n_P}{n_S }\right)}^2 $$ (2)这里,φ P是产品的量子产率,φ S 参考的量子产率。术语slopeS 和slopeP 分别表示从积分荧光强度与参考和产品吸光度的关系图中得出的斜率。 n P 和 n S对应于所用溶剂的折射率[25]。
蛋白质泄漏
对于浸出实验,未稀释的颗粒分散体通过改性聚醚砜膜(MWCO =100 kDa 或 300 kDa,Pall,Dreieich,德国)通过离心(16,000 g,5 分钟)进行超滤。
热稳定性
为了分析热稳定性,纳米粒子和纯 GFP 在 20 或 60°C 下分别保持 0 和 24 小时。如上所述稀释纳米颗粒和纯 GFP。
光漂白
为了研究 GFP 掺杂的纳米粒子和纯 GFP 对光漂白的稳定性,将溶液暴露在七个绿色 LED 发出的光下长达 20 分钟。在 t 采集的样品的荧光强度 =0、2 和 20 分钟被测量。
对蛋白质降解的稳定性
为了确定 GFP 对蛋白酶 K 的稳定性,纯 GFP、未标记的二氧化硅纳米粒子 (CUS1US2U) 与额外的 GFP 和三倍标记的二氧化硅纳米粒子 (CFS1FS2F) 以相同的 GFP 浓度和相同数量的粒子使用。所有样品均按 1:100 稀释。对于 10 个 GFP 分子的数量,选择了一个蛋白酶 K 分子。在添加酶之前,在上述条件下测量了一个样品。添加后,在t之后进行测量 =0、15、30、45、60 和 90 分钟。
细胞摄取实验
为了确定细胞对纳米颗粒和GFP的内化,使用肺癌细胞系A549(ACC-107)进行了细胞摄取实验。
细胞培养
A549 细胞(DSMZ,Braunschweig,德国)在 T75 烧瓶(Greiner bio-one,Frickenhausen,Germany)中使用 Dulbecco's Modified Eagle 培养基(DMEM,Thermo-Fisher-Scientific,Waltham,MA,USA)培养小牛血清 (FCS)。 2 × 10 4 cm −2 A549 细胞接种在 12 孔板的盖玻片上并培养 24 小时。然后用 GFP 掺杂的纳米颗粒和 GFP 溶液在 1 mL 培养基中处理细胞 24 小时。纳米颗粒的 SiO2 浓度为 37 μg mL -1 而 GFP 浓度为 5 μg mL -1 对于纳米颗粒和纯 GFP。处理后,用磷酸盐缓冲液(PBS)洗涤细胞两次。
样品制备和共聚焦成像
细胞在室温下用 4% 多聚甲醛的 PBS 溶液固定 20 分钟。对于细胞膜染色,四甲基罗丹明缀合的 WGA(小麦胚芽凝集素(2 μg mL -1 (在 PBS 中)、W849、Thermo-Fisher-Scientific(Invitrogen),Waltham,MA,USA)并在室温下孵育 10 分钟。用 PBS 洗涤三个步骤后,用 PBS 洗涤细胞三次,并用 Mowiol/DABCO(Carl Roth,Karlsruhe,Germany)固定在载玻片上。
共聚焦图像是在 TCS SP5 系统(Leica,Wetzlar,Germany)上拍摄的。对于成像,63 倍油浸物镜 (n =1.518)。使用氩离子激光线在 λ 处进行连续扫描 =488 nm (25%) 用于激发 GFP 和二极管泵浦固态激光器 λ =561 nm (25%) 用于激发四甲基罗丹明。
结果与讨论
本研究旨在在合适的条件下用 GFP 功能化二氧化硅纳米粒子,以保持蛋白质的生化特性和功能。在之前的工作中,我们在双相环己烷/水系统中使用 ʟ-精氨酸控制的四乙氧基硅烷 (TEOS) 水解,合成了尺寸范围在 15 到 80 nm 之间的近红外染料掺杂的单分散荧光二氧化硅纳米粒子 [26]。在这里,我们采用了这种合成程序,将 GFP 作为模型蛋白嵌入到二氧化硅基质中。在方案 1 中,示意性地描述了粒子合成的程序。 GFP 掺杂和非掺杂结构(核/壳)分别以绿色和灰色突出显示。在第一步中,获得了 GFP 掺杂的二氧化硅核颗粒 (CF)。随后的再生步骤(CFS1 和 CFS1S2)允许合成更大的粒径。在第一个再生步骤中,外壳被修改为(CFS1F)或没有(CFS1U)掺入蛋白质。同样,在第二个再生步骤中,添加了标记的(CFS1FS2F、CFS1US2F)或未标记的(CFS1FS2U、CFS1US2U)外壳。这些变化可以很好地控制嵌入蛋白质的数量及其在指定壳或颗粒核中的定制排列。此外,合成了没有任何嵌入 GFP(CU、CUS1U 和 CUS1US2U)的纯二氧化硅纳米粒子,以研究蛋白质嵌入对粒子特性的潜在影响。此外,对于所有这些步骤,将 GFP 溶解在不同 pH 值的两种不同缓冲系统(ʟ-精氨酸和 NaHCO3)中,以确定蛋白质溶剂对颗粒合成、形态、荧光强度、发射波长和ζ电位。
<图片>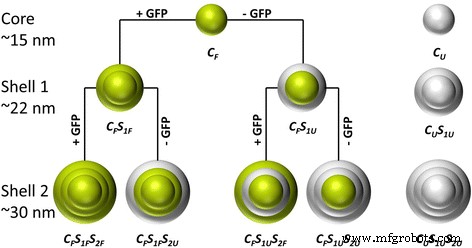
合成粒子及其粒子结构的概述。绿色分别表示 GFP 嵌入到核或壳中。灰色代表没有任何 GFP 的壳(CF =核心荧光,CU =核心未标记,SF =壳荧光,SU =壳未标记,S1 =第一壳层,S2 =第二壳层)
纳米粒子表征
物理粒子属性的确定
为了描述掺入 GFP 后的粒径和形态,并为了确定两种不同缓冲系统对这些特性的影响,记录了 TEM 图像(图 1)。进一步的 GFP(NaHCO3) 改性、GFP(ʟ-精氨酸) 改性和未标记纳米粒子的 TEM 图像显示在 SI 中(附加文件 1:图 S1,附加文件 2:图 S2,附加文件 3:图 S3,附加文件4:图S4)。在具有两个再生长步骤的合成程序之后,获得了三种不同的粒径。核心颗粒的尺寸约为 15 nm,第一次再生步骤后的颗粒尺寸约为 22 nm,第二步后的颗粒尺寸约为 32 nm。总之,所有纳米粒子都近似球形,并表现出窄的尺寸分布(p <10%)。选择三代完全染色的GFP(ʟ-精氨酸)纳米粒子(CF、CFS1F和CFS1FS2F)和GFP(NaHCO3)(CF)核心纳米粒子作为模型。
<图片>
三代 GFP-ʟ-精氨酸修饰纳米颗粒和 GFP(NaHCO3) 修饰纳米颗粒核心颗粒的 TEM 图像。在 a , c 和 d , GFP(ʟ-arginine) 的三代:CF 核心粒子 (a , dTEM =15.5 ± 1.1 nm);第一次再生步骤后的 CFS1F 纳米粒子(核 + 壳 1)(c , dTEM =23.5 ± 2.0 nm) 和 CFS1FS2F 在第二次再生步骤后(核 + 壳 1 + 壳 2)(d , dTEM =35.3 ± 2.0 nm)。在b ,显示了 GFP(NaHCO3) 标记的核心纳米粒子 (dTEM =15.2 显示为
比较不同 GFP 掺杂和未标记纳米粒子的大小(表 1),值得注意的是,相同数量的再生长步骤导致相同的平均粒径,与蛋白质的存在或使用的缓冲溶液无关。未标记的颗粒具有也类似大小(CU:dTEM =13.4±0.4纳米,的DDL =10±3纳米; CUS1U:dTEM =20.9±1.3纳米,的DDL =20±6纳米; CUS1US2U:dTEM =33.2±1.0纳米,DDLS =38 ± 10纳米)。
总之,证明将蛋白质掺入二氧化硅基质和提供蛋白质的缓冲溶液中对所得粒径和形态没有显着影响。
据我们所知,文献中没有描述其他嵌入 GFP 的二氧化硅纳米粒子,它们表现出类似的小尺寸以及同样窄的尺寸分布 (<10%) [20, 27]。这种小纳米粒子在细胞内蛋白质递送以及癌症诊断和治疗领域具有广阔的应用潜力[28]。
ζ-潜力
所有纳米粒子的 ζ 电位是通过使用它们的电泳迁移率计算确定的。所有类型的掺杂纳米粒子都表现出负 ζ 电位,绝对值范围从 - 28 到 - 36 mV(图 2)。相比之下,未标记粒子的 ζ 电位显示出非常相似的值,核心粒子为 − 35.5 ± 2.0 mV,第一次再生步骤后为 − 34.0 ± 3.7 mV,第二步后为 − 34.5 ± 1.2 mV。这些高度负的 ζ 电位 (<− 28 mV) 值表明纳米颗粒对由于静电排斥引起的团聚具有高稳定性。与未标记纳米粒子的 ζ 电位相比,数据表明所得粒子大小和 GFP 掺入粒子核或壳的粒子基质中均未对粒子电荷产生显着影响。
<图片>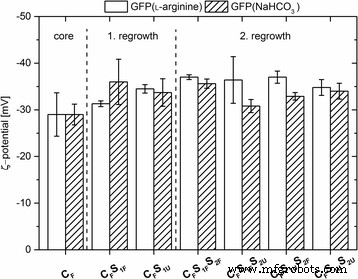
标记纳米粒子的 ζ 电位 [mV]。从溶解在 7.2 mM ʟ-精氨酸或 10 mM NaHCO3 中的 GFP 开始制备纳米颗粒。 误差线 表示三个测量值得出的标准偏差
光谱研究
荧光光谱
所有 GFP 掺杂的二氧化硅纳米粒子都表现出类似的发射最大值 (λ em =508 nm),这也与游离 GFP 的发射最大值相当(SI,附加文件 5:图 S5)。为了比较各种标记纳米粒子的荧光强度,将纳米粒子浓度归一化(SI 5 中的计算)。正如预期的那样,逐步添加标记的壳导致纳米颗粒的荧光增加(图 3)。
<图片>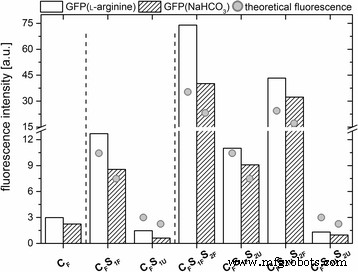
对于各种粒子系统中的每一个,在 508 nm 处发射最大值的归一化荧光强度。此外,理论荧光强度(灰点 )与颗粒体积的增加有关
仅具有标记核但具有非掺杂壳的纳米颗粒表现出最低的荧光。带有一个附加标记外壳的纳米颗粒显示出中等荧光,带有两个标记外壳的纳米颗粒显示出最强的荧光(图 3)。值得注意的是,与具有掺杂外层的纳米颗粒相比,添加非掺杂外壳似乎略微降低了纳米颗粒的荧光。这种效应可能是由未标记的二氧化硅壳的屏蔽效应引起的。总之,在核心粒子中添加 GFP 掺杂的壳导致所得纳米粒子的荧光强度增加,这似乎与伴随纳米粒子生长的体积变化有关。
与通过从溶解在 NaHCO3 中的 GFP 开始的模拟嵌入过程获得的纳米粒子相比,在纯化后最初溶解在 ʟ-精氨酸中的 GFP 嵌入导致所得纳米粒子的荧光强度高 1.3 倍。类似地,与在 NaHCO3 中稀释的 GFP 相比,在 ʟ-精氨酸中稀释的 GFP 表现出更高的荧光强度(附加文件 5:图 S5)。这种影响可能是由缓冲液的不同 pH 值解释的(pHʟ-精氨酸 =10.3,pH\( _{{\mathrm{NaHCO}}_3} \) =9.2)。
出于这个原因,纯 GFP 的荧光被系统地测量为 pH 值的函数(SI,附加文件 6:图 S6)。数据显示随着pH值在pH值5.5 - 10.5范围内增加,荧光呈双曲线形增加。结果与其他关于 GFP 依赖于 pH 的荧光的报告一致。对于野生型 GFP,据报道荧光在 pH 6 - 10 范围内不变,但在较低 pH 下会降低,而在 pH 值> 10 时会增加 [29]。此外,可以通过引入点突变来改变 GFP 的 pH 敏感性 [30]。与 Aequorea 相比,本研究中使用的 GFP 具有三点突变 野生型蛋白,即 S2A、F64L、S65T。其中,当在 480 nm 激发时,第 65 位的丝氨酸被苏氨酸取代会增加蛋白质的荧光强度,因为该氨基酸参与生色团的形成。此外,S65T/F64L 变体表现出 pH 依赖性荧光 [30]。 GFP掺杂的纳米颗粒(CF)表现出类似的pH依赖性荧光(图3),表明pH依赖性机制不受嵌入过程的影响。
荧光量子产率
为了进一步表征荧光纳米粒子的特性,确定了它们的量子产率。这是通过绘制积分荧光强度与 488 nm 处的吸光度来实现的(图 4)。随后,使用方程计算量子产率。 2. 以罗丹明 6G 为参考,掺杂 GFP 的纳米粒子 CFS1F 和纯 GFP 的量子产率确定为 φ\( _{{\mathrm{C}}_{\mathrm{F}}{\mathrm {S}}_{1\mathrm{F}}} \) =0.62 和 φpureGFP =0.38。通过使用 Atto488 作为第二个参考(SI,附加文件 7:图 S7)确认了结果。与纯 GFP 相比,GFP 掺杂纳米粒子的更高量子产率似乎是由 GFP 封装到二氧化硅基质中引起的,并且可能与蛋白质的空间固定或二氧化硅基质提供的化学环境改变有关.
<图片>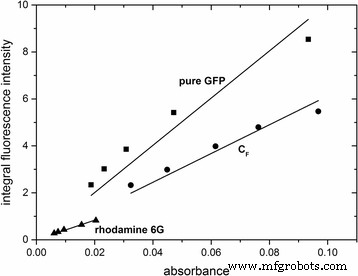
GFP 掺杂颗粒和纯 GFP 的积分荧光强度与 488 nm 处的吸光度图。罗丹明 6G 用作参考。线性相关由直线拟合 .对应的线性方程如下:y pureGFP =1.00554 × 10 10 × x , R 2 =0.97712; y\( _{C_F{S}_{1F}} \) =6.12332 × 10 9 × x , R 2 =0.99331; 是 罗丹明6G =4.1772 × 10 9 × x , R 2 =0.99678
粒子稳定性
蛋白质泄漏
进行浸出实验以证明 GFP 掺杂的纳米粒子的结合稳定性。使用允许 GFP (MW~27 kDa) 通过但保留纳米颗粒的 MWCO 通过膜超滤后,滤液中未检测到荧光,表明 GFP 与二氧化硅基质永久偶联。
分析超速离心
为了支持获得的结果并确定与颗粒基质结合的 GFP 类型,进行了分析超速离心。为此,在相同的粒子和 GFP 浓度下测量了标记的 CFS1FS2F 粒子和未标记的 CUS1US2U 粒子与 GFP 混合。结果(附加文件 8:SI 中的图 S8)表明大多数 GFP 分子在合成过程中嵌入了二氧化硅基质中。
热稳定性
为了确定它们的热稳定性,分别在室温和 60°C 下孵育后测量了 CF 与纯 GFP 相比的荧光信号(图 5)。在室温下 24 小时后,未检测到两个样品的荧光降低,表明对蛋白质稳定性没有影响。然而,在 60°C 的高温下 24 小时后,只能观察到 CF 初始荧光强度的 20%,而纯 GFP 的荧光信号没有恢复。这强烈表明与纯 GFP 相比,嵌入 GFP 的二氧化硅具有更高的热稳定性。 Since an elevated temperature leads to a significant increase in the thermal motions of the protein molecule, which can disrupt its structure, it is hypothesised that the surrounding silica matrix protected the GFP against external influences by spatial constraints.

Influence of temperature (r.t., 60 °C) on the fluorescence of GFP-doped particles (CF, ʟ-arginine) and pure GFP. The normalised fluorescence intensity [%] of the emission maximum at 508 nm versus time [h] is shown
Photostability
Furthermore, the photostability of the samples was tested. For measurements, the nanoparticle stock suspension (CF, ʟ-arginine) was diluted tenfold. Pure GFP was diluted in ʟ-arginine according to the calculated concentration of GFP in the nanoparticle suspension. After exposure of the samples to light of a green LED array over a period of time up to 20 min, the fluorescence intensity was measured (Fig. 6). Within 20 min, the fluorescence intensity of the nanoparticle suspension decreased only slightly. After 20 min, 89% of the initial fluorescence (100%) of the nanoparticles was preserved. In comparison, the pure GFP seemed to be more affected by light exposure. After 20 min, only 81% of the initial fluorescence of pure GFP remained. This result indicated, that GFP, when embedded into silica nanoparticles, was better protected from photochemical alterations induced by the LED light than the pure protein.

Photostability of GFP-doped nanoparticles (CF) and pure GFP in ʟ-arginine. The normalised fluorescence intensity [%] of the emission maximum at 508 nm was measured after exposure to LED light for the given times. Data are mean values. Error bars indicate the standard deviation
Stability Against Protein Degradation
As a further characterisation step, the degradation of GFP in the presence of proteinase K was tested. Therefore, three different systems were used (pure GFP, unlabelled CUS1US2U mixed with GFP and labelled CFS1FS2F). For all systems, equal amounts of GFP and particles were used. After 90 min of incubation, the fluorescence intensity of pure GFP and unlabelled particles with added GFP decreased to 5 - 7% of the initial fluorescence intensity, whereas the one of the labelled particles decreased to 52% (Fig. 7). This result indicates that the GFP is protected by the silica matrix and is degraded slower than free GFP in presence of proteolytic enzymes.

Stability against protein degradation of pure GFP (grey ), unlabelled particles mixed with GFP (CUS1US2U, blue ), and GFP-doped silica nanoparticles (CFS1FS2F, green )。 The normalised fluorescence intensity [%] of the emission maximum at 508 nm was plotted against the incubation time [min] with proteinase K
To conclude, the encapsulation of GFP into silica matrix appeared to bring about significant advantages:The stability of the protein was increased not only against elevated temperatures and light-induced photobleaching but also against the degradation through enzymes. Therefore, the silica matrix seems to protect the embedded GFP as compared to the free GFP.
Cellular Uptake Experiments
In order to determine, if the GFP-doped nanoparticles are able to deliver their cargo into cells, uptake experiments were performed (Fig. 8). A549 cells were exposed to GFP-doped nanoparticles and for comparison to the pure protein. In order to optimise the GFP load of the particles for imaging, a higher amount of GFP as compared to the nanoparticles described before was embedded into the particles. More specifically, a 20-fold amount of GFP in ʟ-arginine was used to label the second shell of the CFS1FS2F particles. These nanoparticles were diluted to a final concentration of 37 μg SiO2 per millilitre in cell culture medium and incubated for 24 h with the cells. The amount of GFP in both samples (nanoparticles and pure GFP) was 5 μg mL −1 .

Confocal microscopy images of A549 cells after 24 h exposure to GFP dissolved in ʟ-arginine (A1 –A3 ) and GFP-doped nanoparticles CFS1FS2F (B1 –B3 ), and control cells (C )。 Top (1):merge-images; middle (2):Cell membrane (WGA):red; bottom (3):GFP, green . Arrows indicate internalised nanoparticles. Contrast and brightness were enhanced by using the ImageJ software
In order to visualise the cells, the cell membrane was labelled, using tetramethylrhodamine-coupled WGA (wheat germ agglutinin). Confocal imaging was used to localise the GFP-doped nanoparticles and the pure GFP in the cells. After exposure of cells to GFP, no signal related to GFP was observed inside the cell bodies (Fig. 8a). Compared to the control cells, no difference in signal intensity of both channels could be observed (Fig. 8c).
In contrast, after exposure of the cells to the GFP-loaded nanoparticles, bright fluorescence signals were detected in the perinuclear region, indicating internalisation of the loaded nanoparticles through endocytosis. The GFP-loaded nanoparticles appeared to be excluded from the nuclear compartment. A second fraction of agglomerated nanoparticles was detected on top of the cell membrane (Fig. 8b).
In conclusion, the GFP-doped nanoparticles are internalised by the cells and are able to transport their cargo into the cells. After exposure of the cells to GFP, fluorescence signals were not detected inside the cell body. This finding is in line with the results of Pesce et al. [31], who did not observe cell-associated fluorescence after incubation of A549 cells with GFP for 24 h. The lack of cell-associated GFP signals might be due to the fact that GFP is not internalised by the cells. Alternatively, GFP fluorescence might be quenched by the low pH value present in endocytic vesicles or lysosomes or degraded by proteolytic enzymes. Therefore, the fluorescence signals of the nanoparticles might indicate a protective effect of the silica nanoparticle matrix against lysosomal degradation.
Conclusions
In this study, a novel approach is presented for synthesis of monodisperse GFP-doped silica nanoparticles with a mean particle-core size of 15 nm. By subsequent growth steps, the particle size and the amount of embedded GFP can be varied. At the end of this procedure, the fluorescence properties of GFP are kept. Incorporation of GFP into additional outer shells results in an increase in the nanoparticle fluorescence. Coverage of the nanoparticles by non-doped shells seems to slightly decrease their fluorescence. These properties indicate the potential to incorporate cargo molecules into specific particle shells. The GFP-doped nanoparticles exhibit a higher quantum yield as compared to the pure GFP. The incorporation into the silica matrix appeared to be durable, as no leaching of protein was detected by ultrafiltration. The silica matrix also seems to improve the thermal properties and photostability of the protein. Furthermore, it is possible to encapsulate different proteins in the different shells, in order to prepare multifunctional nano-carriers. Finally, the nanoparticles are applicable for intracellular delivery of their cargo. The incorporation of proteins into the particle matrix seems to increase delivery and reduce lysosomal degradation of the cargo. Therefore, the protein-doped silica nanoparticles constitute a promising novel tool for biomedical applications of nanoparticles, especially in the field of intracellular delivery of macromolecules.
缩写
- AUC:
-
Analytical ultracentrifuge
- DLS:
-
动态光散射
- FCS:
-
Foetal calf serum
- GFP:
-
Green fluorescent protein
- MW:
-
Molecular weight
- MWCO:
-
Molecular weight cut off membrane
- PBS:
-
Phosphate buffered saline
- r.t.:
-
Room temperature
- TEM:
-
透射电子显微镜
- TEOS:
-
Tetraethoxysilane
纳米材料
- 用于合成和生物医学应用的荧光纳米材料的进展和挑战
- 131I 追踪的 PLGA-脂质纳米颗粒作为靶向化疗治疗黑色素瘤的药物递送载体
- 轻松合成锚定在 MWNT 上的 SiO2@C 纳米粒子作为锂离子电池的高性能阳极材料
- 用于体内 CT 成像和肾脏清除特性的新型生物相容性 Au Nanostars@PEG 纳米颗粒
- 合成单分散二元 FePt-Fe3O4 纳米粒子的后处理方法
- 从豆腐废水中合成荧光碳量子点的简单方法
- 用于光热疗法和光声成像的聚吡咯涂层铁铂纳米粒子的合成和体外性能
- 铜纳米粒子合成和稳定方面的环保能力:催化、抗菌、细胞毒性和抗氧化活性
- 用于氧化还原刺激触发释放的新型双线粒体和 CD44 受体靶向纳米颗粒
- 桔梗皂苷(桔梗)用于金和银纳米颗粒的绿色合成
- 辣根过氧化物酶封装的空心二氧化硅纳米球用于细胞内活性氧种类的传感
- 基于生物矿化合成铈掺杂碳质纳米粒子的高羟基自由基清除活性