

工程纳米粒子对海洋浮游植物中外聚物质释放的影响
摘要
工程纳米粒子 (ENP) 是现代纳米技术的产物,可能会影响海洋环境,从而对海洋生态系统构成严重威胁。然而,海洋浮游植物对 ENPs 的细胞反应仍未完全确定。在这里,我们调查了四种不同的硅藻物种(Odontella mobiliensis , Skeletonema grethae , 三角褐指藻 , Thalassiosira pseudonana ) 和一种绿藻 (Dunaliella tertiolecta ) 在模型 ENP 处理下的细胞外聚合物 (EPS) 释放:25 纳米二氧化钛 (TiO2)、10-20 纳米二氧化硅 (SiO2) 和 15-30 纳米二氧化铈 (CeO2)。我们发现 SiO2 ENP 可以显着刺激这些藻类的 EPS 释放(200-800%),而 TiO2 ENP 暴露引起的释放最低。此外,细胞内Ca 2+ ENPs 可以触发浓度,这表明 EPS 的释放过程是通过 Ca 2+ 介导的 信号通路。随着对ENP诱导EPS释放介导的细胞机制的更好理解,可以制定潜在的预防和安全措施,以减轻对海洋生态系统的负面影响。
背景
尺寸在 1 到 100 nm 之间(至少在一个维度上)的工程纳米粒子 (ENP) 用于制造多种消费品,包括打印机墨水和油漆、洗涤剂、杀菌剂、涂料、化妆品、防晒乳液、轮胎、计算机构造和药物输送。鉴于 ENP 的应用前景广阔,2017 年仅美国国家纳米技术计划 (NNI) 的资金就接近 14 亿美元 [1,2,3]。在纳米尺度上建立基础知识是纳米技术研究界第一阶段的主要重点。截至 2009 年,这一新知识支撑着全球约 1/4 万亿美元的市场,其中约 910 亿美元来自包含纳米级组件的美国产品 [4]。随着纳米技术的飞速发展,ENPs最终会进入水生系统是不可避免的。 ENPs 在环境中的潜在毒性(例如,产生活性氧,ROS 的潜力)方面的主要问题与其巨大且独特的表面反应性有关。然而,由于自然水域复杂的环境和生物因素以及各种ENPs,对海洋生态系统的实际影响在很大程度上仍然未知[1, 5, 6]。先前的研究表明,ENPs 会对以藻类为基础的海洋生态系统造成重大危害 [7, 8]。海洋生物(尤其是浮游植物)已显示与 ENP 相互作用导致负面影响 [9,10,11]。随着纳米技术在各个领域的潜在应用增加,越来越多的 ENP 可能进入水生环境,因此海洋浮游植物对 ENP 的细胞反应值得进一步关注 [12,13,14,15,16,17,18,19,20, 21]。
大多数海洋微生物,无论是自养还是异养,通常都能够产生外聚合物(EPS),其在海洋生态系统中具有多种功能作用和物理特性,可作为生长抑制剂、生长促进剂、毒素、金属清除剂或作为底物异养循环 [22,23,24,25,26]。从海洋中的浮游植物和细菌释放的 EPS 是富含多糖的阴离子胶体生物聚合物,对于海洋凝胶、海洋雪和生物膜的形成以及胶体和微量元素的清除以及提供针对各种环境威胁的保护至关重要,包括 ENP [7, 15, 19, 20, 25, 27]。此外,EPS的分泌被认为是浮游植物受到各种胁迫时的一种自然反应[8]。
Ca 2+ 是参与多种细胞内信号通路的常见第二信使。已经证明 Ca 2+ 是硅藻 Amphora coffeaeformis 的趋化性、运动性和粘附所必需的 [28]。增强的细胞内游离 Ca 2+ 已知水平会导致蛋白激酶 C 的激活,这涉及许多细胞内信号通路 [29]。由于EPS的释放与硅藻的运动性和粘附性密切相关,有人提出Ca 2+ -介导的分泌过程控制着硅藻中EPS的释放[30],以及验证Ca 2+ 的直接证据 信号传导、胞吐作用和相关 Ca 2+ 我们之前的研究报道了胞吐作用的信号传导 [31]。过去的研究还表明,与 ENP 的相互作用可以改变细胞内 Ca 2+ 通路,这对细胞信号传导至关重要 [29, 32,33,34]。特定细胞内Ca 2+ 浓度变化在细胞信号传导和分泌过程中很重要;然而,没有关于二氧化钛 (TiO2)、二氧化硅 (SiO2) 或二氧化铈 (CeO2) 改变细胞内 Ca 2+ 的报道 浮游植物水平。
2013 年,Quigg 等人。 [8]总结了ENPs对藻类的直接和间接毒性作用。在我们之前的实验中,ENP 被证明可以促进 EPS 聚合 [35]。在这方面,EPS 可能会加剧或减少 ENP 对水生生物的直接毒性 [7, 15, 36]。然而,从未报道过在 ENPs 胁迫下直接测量浮游植物的 EPS 释放。本研究的目的是研究四种不同硅藻(Odontella mobiliensis , Skeletonema grethae , 三角褐指藻 , Thalassiosira pseudonana ) 和一种绿藻 (Dunaliella tertiolecta ) 在 ENP 处理下。通过了解 ENP 诱导浮游植物 EPS 释放的潜在机制,实施预防和安全措施可以减轻对海洋生物的潜在不利影响。
结果和讨论
ENP 表征
动态激光散射 (DLS) 用于表征以下悬浮在纯水中的 ENP 的尺寸指标:TiO2、SiO2 和 CeO2。 TiO2 的粒径分布范围为 7 至 66 nm,SiO2 的粒径分布范围为 9 至 66 nm,CeO2 的粒径分布范围为 12 至 70 nm。一些较大的尺寸可能是由于聚集或附聚,而 TiO2 的主要尺寸为 25 nm,SiO2 为 10 至 20 nm,CeO2 为 15 至 30 nm,这与制造商的信息一致(图 1)。
<图片>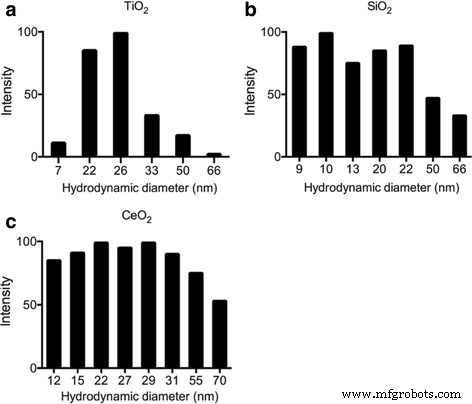
通过 a 的 DLS 评估来表征 ENP 二氧化钛,b SiO2 和 c 超声处理后 L1 培养基中的 CeO2 显示了它们的尺寸分布。 DLS 样品中 ENP 终浓度为 1 μg/ml,超声处理后测量时间为 3 min
ENPs 诱导细胞内 Ca 2+ 浮游植物浓度
探讨ENPs是否能诱导细胞内Ca 2+ 增加 浓度,浮游植物细胞(OD 600 =0.8)加载 Fluo-4AM 染料,并分别暴露于 1 mg/ml 的 25 nm TiO2、10-20 nm SiO2 和 15-30 nm CeO2 ENP。细胞内Ca 2+ 的变化 以浮游植物细胞内的荧光强度表示的浓度监测 150 秒。图 2a-e 显示 1 mg/ml 的三种 ENP 增加了 Ca 2+ SiO2 中的浓度约 50-300%,TiO2 约 40%,CeO2 约 150-200%,而控制条件(L1 介质)保持不变。结果表明ENPs可诱导显着的细胞内Ca 2+ 浮游植物中的反应并表明浮游植物通过 Ca 2+ 对不同的 ENPs 作出反应 信号通路。我们的数据表明细胞内 Ca 2+ 的变化很小 TiO2 存在时的水平,这可能归因于 TiO2 诱导的毒性引起的大量浮游植物细胞死亡 [37, 38]。在我们之前的研究中,TiO2 促使细胞内 Ca 2+ 增加 浓度 [34] 以及显着的细胞凋亡 [39]。然而,SiO2 令人惊讶地表现出最明显的细胞内 Ca 2+ 所有浮游植物物种的增加,而 CeO2 只能触发中间细胞内 Ca 2+ 浓度增加。先前的研究表明,高浓度的 CeO2 (> 50 mg/ml) 有可能诱导细胞内氧化应激和细胞内 Ca 2+ 尽管影响很小,但支持我们的发现 [40]。我们还测量了人工海水中每个ENPs的zeta电位,以解决表面电荷可能引起的潜在影响;然而,价值很低。测量结果表明 ENP 被认为近似中性 [41](附加文件 1:补充数据)。这是第一个报告,其中发现不同的 ENPs 诱导细胞内 Ca 2+ 特定浮游植物的浓度变化,最终为未来的研究铺平了新的道路。
<图片>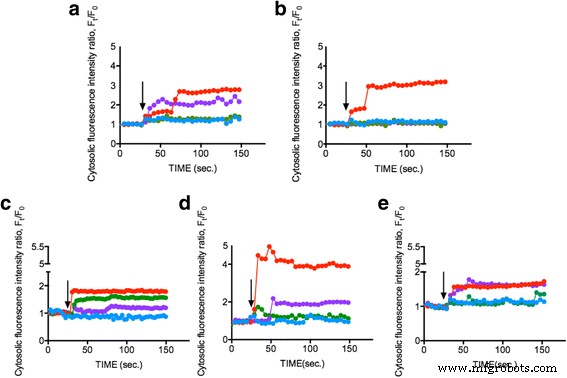
细胞内Ca 2+ 的测定 不同ENPs刺激后的浓度。不同的浮游植物细胞a 杜氏杜氏藻 , b Thalassiosira pseudonana , c 骷髅草 , d 三角褐指藻 , 和 e 运动齿状藻 用浓度为 1 mg/ml 的 TiO2 25 nm(绿色)、SiO2 10-20 nm(红色)、CeO2 15-30 nm(紫色)和对照(蓝色)处理。黑色箭头表示应用 EPN 的时间点(30 秒)。测量显示来自平均 20 个单个细胞的代表性数据
ENP 诱导的浮游植物 EPS 释放
酶联凝集素测定 (ELLA) 用于评估当用 TiO2、SiO2 和 CeO2 ENP 刺激时浮游植物细胞释放的 EPS 量,浓度范围为 1 μg/ml 至 5 mg/ml,基于先前对 TiO2 的研究[ 42, 43] 和 CeO2 [44,45,46]。 EPS 分泌被标准化为总浮游植物 DNA 量(附加文件 1:补充数据),以便具有相同的比较基础。与对照相比,我们发现 10-20 nm SiO2 能够将 杜氏藻 中的 EPS 释放量提高多达 550% , 500% Thalassiosira , 骨架 1000% , 400% Odontella 和 900% 在 褐指藻 (图3)。当浮游植物物种暴露于 TiO2 时,对 EPS 分泌没有强烈影响,因为只有 Skeletonema 和 褐指藻 表现出明显的变化。 EPS 释放数据因此与我们的细胞内 Ca 2+ 一致 浓度结果。 TiO2对EPS的产生没有显着影响,类似于细胞内Ca +2 由于二氧化钛对浮游植物的毒性,浓度显示出非常有限的变化。 ROS 的产生和残留会导致许多并发症,例如浮游植物的细胞凋亡 [47,48,49]。在 CeO2 处理中,结果显示对 杜氏藻 的影响很小 , 骨架 , Odontella , 和 褐指藻 .然而,SiO2在Thalassiosira pseudonana中表现出最显着的EPS诱导 (大约 600%)和 Skeletonema grethae (大约 1000-1500%)。这些数据表明不同的ENPs可以诱导浮游植物和细胞内Ca 2+ 的特定EPS释放 更改也与 EPS 发布结果相匹配。通过评估细胞内Ca 2+ 的变化 浓度,显然 Ca 2+ 中存在直接联系 ENPs 诱发浮游植物 EPS 分泌的细胞途径。这里的观察结果与我们之前基于 Phaeocystis 的研究一致 EPS 版本 [31]。结果提供了直接证据表明浮游植物可以检测和区分由Ca 2+ 调控的不同EPS释放响应的ENPs 细胞通路。
ELLA 的使用使我们能够通过浮游植物与 ENP 的相互作用来确定 EPS 的释放。我们的研究结果表明,当浮游植物与 SiO2 相互作用时,Dunaliella tertiolecta 的 EPS 分泌显着增加 , Thalassiosira pseudonana , 和 Skeletonema grethae .看来这些硅藻已准备好识别 SiO2 颗粒。然而,在三角褐指藻 , 没有发现强烈的 EPS 分泌。这种差异代表由 ENP 触发的 EPS 释放取决于浮游植物种类和 ENP 浓度(图 3)。在之前的一项研究中,石油泄漏导致大量海洋微生物 EPS 释放,被提议用来抵消石油泄漏的负面后果 [50]。此外,Boglaienko 和 Tansel 发现 SiO2 颗粒能够有效去除油聚集体 [51]。我们的发现提供了一种新的潜在机制,其中低毒性 SiO2 颗粒可以诱导特定浮游植物释放 EPS,通过促进 EPS 聚集潜在地促进溢油清除。从未有报道称二氧化铈会干扰以浮游植物为基础的海洋生态系统。这里的结果表明,除了 Thalassiosira pseudonana 外,CeO2 ENP 可以影响这里的所有浮游植物。 CeO2 ENPs 可能与 SiO2 一样,具有促进特定浮游植物 EPS 释放的能力,用于石油减排应用。
结论
由于当前和未来纳米材料的排放,ENP-海洋环境相互作用变得越来越重要。在这里,我们证明了增强的 EPS 分泌是 ENP 对浮游植物的主要影响之一。我们还提供证据表明,不同的浮游植物可以通过调节 Ca 2+ 对各种 ENP 胁迫做出不同的反应。 途径。然而,完整的ENPs对海洋生态系统的评估需要进一步的研究,以提供对纳米材料与海洋生物之间相互作用的详细知识和理解。
<图片>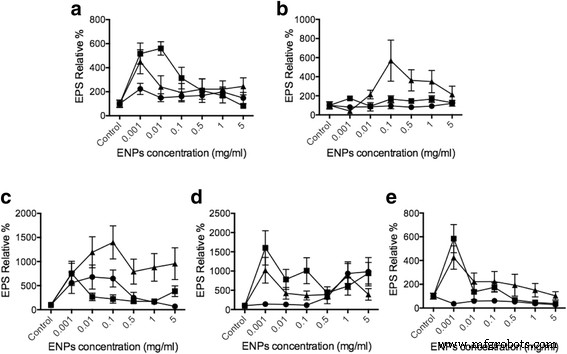
由各种 ENP 触发的 EPS 释放。不同的浮游植物细胞a 杜氏杜氏藻 , b Thalassiosira pseudonana , c 骷髅草 , d 三角褐指藻 , 和 e 运动齿状藻 分别用 TiO2(圆圈)、SiO2(三角形)、CeO2(正方形)处理,浓度分别为 5mg/ml 和 1mg/ml、0.5mg/ml、0.1mg/ml、10μg/ml、1μg /ml (n =3)
方法
浮游植物培养
Odontella mobiliensis的分批培养 (CCMP597), Dunaliella tertiolecta (UTEX999), Skeletonema grethae (CCMP775), 三角褐指藻 (UTEX646), Thalassiosira pseudonana (Provasoli - Guillard 海洋浮游植物培养物保藏中心,美国明尼苏达州西布斯贝港)在 L1 海洋培养基(Sigma,MO,美国)中以 100 μmol m -2 的 14:10(亮:暗)循环生长 s −1 和 24°C 在无菌条件下。用血细胞计数器进行细胞计数确定培养物的生长期。
纳米粒子和表征
所有 ENP、TiO2、SiO2、CeO2(Sigma-Aldrich,MO,美国)在使用前都在纯水中进行了超声处理。在测试之前,ENP 用过滤的 L1 培养基(Sigma,MO,USA)重构。使用零差动力学激光散射 (DLS) 独立确认 ENP 的大小。简而言之,将海水样品通过 0.22-μm Millipore 膜(用 0.1N HCl 预洗)重新过滤并直接倒入五个 10ml 散射池中,然后将其放置在 Brookhaven BI-200SM 激光光谱仪(Brookhaven Instruments,纽约,美国)。在45°角检测到的散射强度波动的自相关函数由Brookhaven BI 9000AT自相关器在线处理,并通过CONTIN方法(Provencher,1982)计算粒度分布。超声处理后立即收集每个样品的结果,一式三份。使用单分散乳胶微球的标准悬浮液(Polysciences,PA,USA)对 DLS 光谱仪进行校准。
ENP 治疗
浮游植物细胞在具有 L1 培养基的 96 孔板中培养 24 小时。细胞用 ENP 原液处理:5 毫克/毫升和 1 毫克/毫升、0.5 毫克/毫升、0.1 毫克/毫升、10 微克/毫升、1 微克/毫升的 TiO2、SiO2 和 CeO2(Sigma-Aldrich,MO ,美国)或 L1 培养基(对照)48 小时。收集含有分泌型 EPS 的上清液并在 4000 rpm 下短暂离心以去除残留的 ENP。该协议改编自我们之前的出版物 [34]。这里使用的浓度范围并不是为了代表或模拟环境中当前的 ENP 水平,而是旨在评估 ENP 对海洋浮游植物的全部潜在影响并研究相关的细胞机制。作为一种有前途的新兴纳米材料,ENPs 尚未达到其全部商业能力。在ENPs进入商业和家庭产品市场以将更多ENPs引入海洋之前,迫切需要对其完整的生态影响进行详细评估。
酶联凝集素检测 (ELLA)
收集含有分泌多糖的上清液并在 1700 rcf (Megafuge 1.0R) 下短暂离心以去除残留的 ENP。然后将上清液在 96 孔 (Nunc MaxiSorp, VWR, CA, USA) 板中在 4°C 下孵育过夜。然后用PBST(PBS + 0.05% Tween-20)和PBS洗涤96孔板,然后用1% BSA封闭。 96 孔板再次用 PBST 和 PBS 洗涤,并与凝集素(伴刀豆球蛋白 A,ConA)(Sigma-Aldrich,MO,USA)一起孵育,结合辣根过氧化物酶(HRP;5mg/ml)(Sigma-Aldrich,MO ,美国),在 37°C 下保持 1 小时。在室温下将底物 3,39,5,59-四甲基联苯胺(TMB;Sigma-Aldrich,MO,USA)加入每个孔中,然后加入 H2SO4(Sigma-Aldrich,MO,USA)以终止反应.光密度由 PerkinElmer VICTOR3 (MA, USA) 在 450 nm 处测量。该协议改编自我们之前的出版物 [34, 52]。
DNA 测定
收集含有浮游植物的颗粒并获得 ZR-96 Quick-gDNA 试剂盒(ZYMO Research,CA,USA)。简而言之,4×裂解缓冲液用于破碎浮游植物细胞并流过DNA结合柱,最终被洗脱缓冲液洗脱。 DNA 浓度通过 NanoDrop ND-1000 (Thermo, CA, USA) 测量。方案改编自制造的试剂盒方案。
细胞内 Ca 2+ 的测量 ENPs 引起的浓度
然后用 Fluo-4AM 染料 (1 mM)(Kd =335 nM,λEx =494 nm,和 λEm =506 nm,ThermoFisher,CA,USA)加载浮游植物细胞 60 分钟 [31]。染料加载后,漂洗浮游植物细胞,用 L1 培养基培养,并分别用 1 mg/ml TiO2、SiO2 和 CeO2 处理。所有钙信号实验均在尼康显微镜(Nikon Eclipse TE2000-U,东京,日本)上进行。协议和条件改编自以前的出版物[31, 34]。
ENP 测量的 Zeta 电位
为了测量 ENP 的表面电荷,在 25 °C 的人工海水中,使用 Zetasizer Nano ZS,Malvern 测量 ENP 的 zeta 电位 (ζ)。对每个样品采集数据后,对记录值取平均值。
统计分析
数据报告为平均值±SD。每个实验至少独立进行 3 次。直方图由 GraphPad Prism 6.0 制作。 (GraphPad Software, Inc.,美国加利福尼亚州圣地亚哥)。
纳米材料
- 半导体纳米粒子
- 等离子纳米粒子
- pH 值对花园中黄色染料的影响
- 从气相沉积在无模板表面上的有序密堆积纳米粒子超晶格的形成机制
- 纳米颗粒作为外排泵和生物膜抑制剂,恢复常规抗生素的杀菌效果
- 以叶酸受体为靶点的生物类黄酮染料木素负载壳聚糖纳米颗粒可增强对宫颈癌的抗癌作用
- 纳米技术:从体内成像系统到受控给药
- La1 − xSr x MnO3 锰酸盐纳米颗粒的合成方法对其性能的影响
- PEG 包覆的 CoFe2O4 纳米颗粒的毒性与姜黄素的处理效果
- 酸性胶溶剂对 TiO2 纳米颗粒锐钛矿-金红石比和光催化性能的影响
- 用于氧化还原刺激触发释放的新型双线粒体和 CD44 受体靶向纳米颗粒
- 桔梗皂苷(桔梗)用于金和银纳米颗粒的绿色合成