

M1 巨噬细胞衍生的外泌体 MicroRNA-326 通过介导 NF-κB 信号通路抑制肝细胞癌细胞进展
摘要
越来越多的证据表明,源自 M1 巨噬细胞外泌体的 microRNA (miR) 可以调节肝细胞癌 (HCC) 的进展。然而,尚未报道源自 M1 巨噬细胞外泌体的 miR-326 对 HCC 的影响。因此,本研究的目的是探索来自 M1 巨噬细胞的外泌体 miR-326 调节 HCC 细胞进展的机制。 RT-qPCR 检测到 miR-326 在 HCC 细胞系中的表达。转染改变HCC中miR-326的表达,检测miR-326对CD206和NF-κB表达、细胞增殖、集落形成、迁移、凋亡和侵袭的影响。随后,外泌体从 M1 巨噬细胞中分离出来。 RT-qPCR 鉴定了 M1 巨噬细胞衍生的外泌体中 miR-326 的表达。 M1 巨噬细胞衍生的外泌体中 miR-326 的表达因转染而改变。 M1 巨噬细胞衍生的外泌体与 HCC 细胞共培养,以确定它们对 HCC 细胞生物学进展的影响。最后,进行体内实验以验证体外结果。 MiR-326 在 HCC 细胞中减少,并在 M1 巨噬细胞衍生的外泌体中富集。上调 miR-326 将抑制 HCC 细胞增殖、集落形成、迁移、侵袭和 CD206 和 NF-κB 的表达并促进细胞凋亡,并抑制 HCC 肿瘤体内的生长 ,而下调 miR-326 显示出相反的效果。 M1 巨噬细胞衍生的外泌体抑制 HCC 细胞增殖、集落形成、迁移、侵袭和 CD206 和 NF-κB 表达并增强细胞凋亡,而 miR-326 的过表达增强了 M1 巨噬细胞衍生的外泌体对 HCC 细胞的影响。结果表明,M1巨噬细胞来源的外泌体miR-326通过下调NF-κB的表达抑制HCC的增殖、迁移和侵袭,促进细胞凋亡。
介绍
肝细胞癌(HCC)是全球第五大最常见的癌症,也是最常见的原发性肝癌[1]。中国国家癌症登记中心的数据表明,原发性肝癌的死亡率在常见恶性肿瘤中位列第三,发病率位居第四[2]。 HCC 的主要危险因素是慢性感染丙型肝炎病毒和乙型肝炎病毒、受黄曲霉毒素污染的食品、肥胖、吸烟、大量饮酒和 2 型糖尿病 [3]。经动脉化疗栓塞是中期 HCC 的成熟治疗方法,可提高大多数中期或晚期 HCC 患者的生存率 [4]。目前,HCC的诊断主要依赖于血清生物标志物和影像学技术[5]。 HCC相关的5年生存率仅为60%左右,且近年来发病率逐年升高[6]。因此,研究准确的治疗靶点是HCC治疗的重点。
巨噬细胞是免疫系统的效应细胞和主要调节剂,在组织重塑和修复方面发挥着巨大的作用,并在体内几乎所有组织中执行协调代谢功能[7]。研究表明,M1 巨噬细胞发挥促癌作用并增强 HCC 细胞的运动性 [8]。外泌体是直径为 40-150 nm 的盘状囊泡 [9]。根据Xu等人的研究,外泌体微RNA(miRNA)通过调节靶细胞中的基因表达,对HCC的增殖、侵袭、转移和耐药性发挥作用[10]。一项研究表明,含有外泌体的 miR-326 可能是多发性硬化症的潜在临床靶点 [11]。 MiRNA 可以通过特定序列中的 mRNA 降解和翻译阻断来调节大量蛋白质编码基因的表达,从而充当癌基因或肿瘤抑制剂 [12]。一项研究讨论了 miR-326 通过干扰细胞周期进程和增强细胞凋亡来抑制 HCC 细胞生长,此外还通过降低上皮 - 间质转化表型来抑制细胞侵袭 [13]。另一项研究报道,miR-326 可能是 HCC 患者治疗的潜在治疗靶点 [14]。因此,本研究探讨了外泌体miR-326调控HCC细胞侵袭和迁移的机制。
材料和方法
道德声明
所有动物实验均符合国际委员会的实验动物护理和使用指南。该方案经吉林大学第三医院机构动物护理使用委员会批准。
巨噬细胞的诱导和鉴定
人单核细胞系 THP-1(中国科学院昆明动物研究所,中国昆明)在含有 10% 热灭活胎牛血清的 RPMI 1640 培养基(Gibco,CA,美国;Thermo Fisher Scientific,MA,USA)中培养( FBS)。 THP-1 细胞与 150 ng/mL 佛波醇 12-肉豆蔻酸酯 13-乙酸酯(PMA;P8139,Sigma-Aldrich,SF,CA,USA)反应并在 RPMI 培养基中培养 24 小时以获得 M0 巨噬细胞。然后,通过赖特染色观察诱导前后的细胞形态。 THP-1 细胞和诱导的巨噬细胞重悬于 5 μL PBS 中,滴在载玻片上,用 Wright's 染料溶液染色并与缓冲溶液按 1:2 混合。染色10 min,流水漂洗,显微镜下观察细胞。此外,通过逆转录定量聚合酶链反应 (RT-qPCR) 测量 M0 巨噬细胞标记物(CD68 和 CD206)。接下来,通过与 20 ng/mL 干扰素 (IFN)-γ(#285-IF;R&D Systems,MN,USA)和 10 pg/mL LPS(#8630;Sigma-Aldrich)一起孵育 18 天,将巨噬细胞诱导为 M1 巨噬细胞。 H。 M1巨噬细胞标志物(IDO1和IL-12 p35)通过RT-qPCR检测[15]。
外泌体的提取
通过外泌体分离试剂盒(ExoEasy Maxi Kit,Qiagen,Hilden,Germany)提取外泌体。在无菌条件下将巨噬细胞上清液聚集到 15 mL 离心管中,并通过 0.8 μM 滤膜过滤。各组细胞上清液加入XBP缓冲液(1:1),用exoEasy膜亲和离心柱500g离心 .细胞加入 10 mL XWP 缓冲液并用 3000–5000g 离心 . exoEasy 膜亲和离心柱用 400 μL XE 洗脱缓冲液孵化,500g 离心 .洗脱缓冲液转移至exoEasy膜亲和离心柱,500g离心 .洗脱缓冲液在 4°C 下保存 24 h,然后用于鉴定。其余保存于 − 80 °C。
TEM 观察和纳米粒子追踪分析 (NTA)
将上述得到的外泌体滴入碳载膜铜网中,加入2%磷钨酸。样品在透射电镜下观察,并收集和分析最佳图像。
PBS 中的杂质和颗粒通过 0.22-μM 微孔过滤器去除。根据外泌体的颗粒密度,将过滤后的PBS稀释至合适的浓度,使用Nanosight NS300纳米颗粒检测器(Malvern, Westborough, MA, USA)进行检测。
鉴定后,通过外泌体分离试剂盒(Invitrogen)提取转染miR-326抑制剂和miR-326抑制剂阴性对照(NC)、miR-326模拟物和miR-326模拟物NC的巨噬细胞来源的外泌体。
RT-qPCR
用 Trizol Reagent (Thermo Fisher) 提取总 RNA,并使用 SYBR-Green PCR Master Mix (Roche) 和 ABI 7500 Real-Time PCR System (Life Technologies, Grand Island, NY, USA) 进行实时 PCR )。引物序列见表1。定量分析采用2 -△△Ct 法 .
Western Blot 分析
提取细胞和外泌体的总蛋白。通过二辛可宁酸(BCA)试剂盒(博斯特生物科技有限公司,武汉,湖北,中国)测定蛋白质浓度。蛋白质加入样品缓冲液并在 95 °C 下煮沸,每孔上样 30 µg。蛋白质用 10% 聚丙烯酰胺凝胶电泳 (Boster Biological Technology) 分离并电印迹到聚偏二氟乙烯膜上,然后用 5% 牛血清白蛋白 (BSA) 密封。用针对 CD63 的一抗 (1:1000, Developmental Studies Hybridoma Bank, University of Iowa, Ames, IA, USA)、CD181 (1:1000, R&D Systems)、GAPDH (1:2000, Jackson ImmunoResearch Laboratories, PA, USA) 和用辣根过氧化物酶标记的二抗 (1:500, Jackson ImmunoResearch Laboratories)。图像由Odyssey双色红外荧光扫描成像系统获取,波段灰度值由Quantity One图像分析软件测量。
细胞培养和筛选
选择正常肝细胞系 HL-7702 和人 HCC 细胞系 BEL-7404、HepG2、SMMC-7721 和 QGY-7703 并在含有 10% 胎牛血清 (FBS)、青霉素 (100 U/mL) 的 Gibco RPMI 培养基 1640 中培养) 和链霉素 (100 毫克/毫升)。 RT-qPCR检测MiR-326表达,筛选合适的细胞系。
外泌体标记和外泌体摄取
外泌体用 250 μL Diluent C 重悬,轻轻研磨,避免损坏外泌体膜。将 PKH67 染料(1 μL,Sigma-Aldrich)添加到 250 μL 稀释剂 C 中以达到 500 μL 并孵育。溶液中加入 500 μL 1% BSA,37 °C 孵育 1 分钟。外泌体通过 120,000g 离心获得 4°C 2 小时。 120,000g离心得到PKH67标记的外泌体 4°C 2 小时。外泌体用 6 mL RPMI-1640 培养基重新悬浮,避光。然后,标记的外泌体与 HCC 细胞共培养 12 小时。之后,去除培养基并用PBS洗涤3次,5分钟/次,彻底洗掉未被HCC细胞内吸收的荧光标记的外泌体。外泌体用 4% 多聚甲醛固定,并用 4'-6-二脒基-2-苯基吲哚染色。封口后用激光共聚焦显微镜观察荧光分布。
细胞分组和处理
HepG2细胞和SMMC-7721细胞以0.5-1 × 10 6 接种于12孔板 细胞/孔。在 50-60% 汇合时,用 Lipofectamine 2000 (Invitrogen, Carlsbad, CA) 转染细胞。 HepG2细胞分为miR-326模拟组(转染miR-326模拟物)和NC模拟组(转染miR-326模拟物NC)。 SMMC-7721细胞被分为miR-326抑制剂组(转染miR-326抑制剂)和NC抑制剂组(转染miR-326抑制剂NC)。将 miR-326-mimic、miR-326-inhibitor 及其 NCs 与 Lipofectamine 2000 混合进行转染。未进行任何处理的HepG2细胞和SMMC-7721细胞作为空白组。 miR-326-mimic、miR-326-inhibitor及其NC由广州瑞比生物有限公司(中国广州)设计和组成(表1)。
M1 巨噬细胞衍生的外泌体与 HCC 细胞的共培养
用BCA法检测M1巨噬细胞来源的外泌体悬浮液的蛋白质浓度,并计算出相应的50 μg蛋白质的外泌体悬浮液体积。 HepG2细胞和SMMC-7721细胞以1 × 10 5 接种于12孔板中 每孔细胞/毫升。 HepG2细胞分为4组:对照组(HepG2细胞未与外泌体共培养)、外泌体(Exo)组(HepG2细胞与M1巨噬细胞来源的外泌体共培养)、Exo-miR-326-模拟组(HepG2与转染 miR-326 模拟物的 M1 巨噬细胞衍生的外泌体共培养的细胞,Exo-NC 模拟组(与转染 miR-326 模拟物 NC 的 M1 巨噬细胞衍生的外泌体共培养的 HepG2 细胞)。 SMMC-7721细胞也被分为4组:空白组(SMMC-7721细胞未与外泌体共培养)、Exo组(SMMC-7721细胞与M1巨噬细胞来源的外泌体共培养)、Exo-miR-326-抑制剂组(SMMC-7721 细胞与转染 miR-326 抑制剂的 M1 巨噬细胞衍生的外泌体共培养),Exo-NC-抑制剂组(SMMC-7721 细胞与 M1 巨噬细胞衍生的外泌体共培养,转染了 miR- 326抑制剂NC).
3-(4, 5-Dimethylthiazol-2-yl)-2, 5-Diphenyltetrazolium Bromide (MTT) 检测
用胰蛋白酶分离细胞,接种于96孔板,细胞密度为4 × 10 4 每孔细胞。分别培养12、24、36、48、60 h后弃培养基。用 500 μL 0.5 g/L MTT 溶液孵育细胞,加入 200 μL 二甲亚砜溶液,研磨并孵化。光密度(OD,490 nm)值由酶标仪测定。
菌落形成分析
培养 24 小时并用胰蛋白酶分离,将细胞接种在 35 毫米的小培养皿中,每培养皿有 300 个细胞。每 3 d 更换一次溶液。培养10 d后,用40 g/L -1 固定细胞 多聚甲醛并用 1 g/L −1 染色 结晶紫溶液,干燥。显微镜下计算菌落数(50个以上细胞)。
Transwell 检测
细胞 (1 × 10 5 ) 悬浮于 200 μL 空白培养基中。实验按照Transwell小室(Corning Glass Works,Corning,N.Y.,USA)的说明进行(侵袭实验需要基质胶,但迁移实验不需要)。将 RPIM 1640(10% FBS,600 μL)加入下室。上下室由预涂有基质胶(BD Biosciences,Franklin Lakes,NJ,USA)的Transwell膜隔开。培养24 h,用95%酒精固定。结晶紫染色后,显微镜下5个视野观察细胞。
流式细胞术
细胞周期:细胞被胰蛋白酶分离。细胞 (1 × 10 6 ) 用 0.5 mL PBS 悬浮并研磨成单一悬浮液。与 4.5 mL 预冷的 70% 乙醇在冰上混合,将细胞以 3000g 离心 , 用 5 mL PBS 冲洗并再次以 3000g 离心 .然后,将细胞悬浮在含有 0.2 mg RnaseA 的 1 mL PI/Triton X-100 染色溶液(20 μg PI/0.1% Triton X-100)中。流式细胞仪检测细胞周期。
细胞凋亡:胰蛋白酶消化细胞(1 × 10 6 ) 用 1 mL PBS 悬浮,研磨并以 3000g 离心 .细胞依次用孵育缓冲液(10 mmol/L Hepes/NaOH,pH 7.4,140 mmol/L NaCl,5 mmol/L CaCl2)冲洗,并以3000g离心 .接下来,将细胞与 100 μL 标记溶液(将 FITC-Annexin V 和 PI 添加到孵育缓冲液中以达到 1 μg/mL)一起孵育,以 3000g 离心 ,用孵育缓冲液洗涤一次并用荧光 (SA-FLOUS) 溶液孵化。通过流式细胞术检测细胞凋亡。流式细胞仪检测波长为488 nm,515 nm带滤光片检测FITC荧光,波长大于560 nm的PI检测。结果由计算机自动分析。
裸鼠肿瘤异种移植
将 40 只 4-6 周龄的小鼠(吉林大学动物科学学院,吉林,中国)随机分为 8 组,每组 5 只。小鼠在特定的无病原体级动物实验室饲养1 w,并及时更换饲料、垫子和水瓶。应每天观察小鼠的健康状况。 1周后,将HCC细胞制成细胞悬液,以0.1 mL细胞悬液(1 × 10 6 )。 3-5 d后观察肿瘤生长。裸鼠每4 d称重一次,测量肿瘤体积。注射后20 d处死裸鼠。
统计分析
所有数据均由 SPSS 17.0 软件(SPSS Statistics,Chicago,IL,USA)解释。测量数据表示为平均值 ± 标准偏差。两组之间的比较由t制定 -test,而多组之间的比较通过单向方差分析 (ANOVA) 进行评估。 P 值 <0.05表示差异有统计学意义。
结果
M1 巨噬细胞和外泌体的鉴定
Wright染色法观察PMA诱导的THP-1细胞形态。可见诱导前THP-1细胞体积较小,核质比例较高;诱导后细胞形态不规则,体积变大,核质比例减少;细胞质较丰富,呈淡蓝色,颗粒丰富,有少量液泡;细胞核呈紫红色,常向一侧倾斜,表明细胞具有典型的巨噬细胞形态特征(图1a)。
<图片>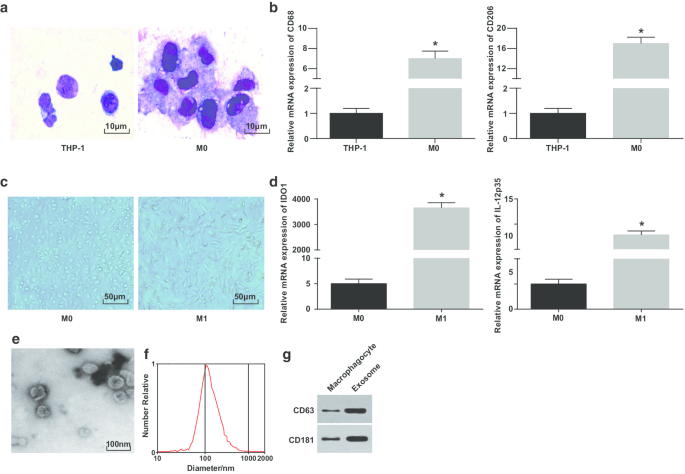
M1巨噬细胞和外泌体的鉴定。 一 赖特染色用于观察 PMA 处理前后 THP-1 细胞的形态。 b RT-qPCR 检测 THP-1 细胞和 PMA 处理的 THP-1 细胞中 CD68 和 CD206 的表达; c 巨噬细胞和 M1 巨噬细胞的形态。 d 通过 RT-qPCR 检测 M0 巨噬细胞和 LPS 和 INF-γ 处理的 M0 巨噬细胞中 IDO1 和 IL-12 p35 的表达。 e 用于外泌体观察的 TEM。 f 通过 NTA 检测外泌体的粒径分布。 g CD63 和 CD181 的蛋白条带。在面板 b 中,*P <0.05 与 THP-1 细胞相比;在面板 d 中,*P <0.05 相对于 M0 巨噬细胞。测量数据被描述为平均值 ± 标准偏差(N =3),两组比较采用t检验
为了进一步验证巨噬细胞的成功诱导,通过 RT-qPCR 测试了诱导前后 CD68 和 CD206 的表达。表明自PMA诱导后CD68和CD206表达升高,表明PMA成功将THP-1细胞诱导为M0巨噬细胞(图1b)。然后,通过诱导 LPS 和 INF-γ,M0 巨噬细胞被极化为 M1 巨噬细胞。观察和测试巨噬细胞的形态和M1型巨噬细胞表面标志物IDO1和IL-12 p35的表达。 M0型巨噬细胞呈各种不规则粘附形态,呈圆形、椭圆形或纺锤形。由 IFN-γ 刺激的巨噬细胞的形态表现出更多的伪足和突起和梭形(图 1c)。 RT-qPCR概述(图1d)LPS和INF-γ处理后,M1巨噬细胞的标志物(IDO1和IL-12p35)增加。
随后,通过 TEM 观察到来自巨噬细胞的外泌体。结果表明,巨噬细胞来源的外泌体丰富,形状呈圆形或椭圆形,膜状结构,大小均匀,污染物较少(图1e)。 NTA 显示具有集中 MODE 曲线峰且线性平滑的外泌体具有更集中的直径和更少的杂质(图 1f)。蛋白质印迹分析报告说,与巨噬细胞相比,来自巨噬细胞的外泌体中特异性标记蛋白 CD63 和 CD181 的表达升高(图 1g)。这些结果表明我们成功地诱导单核细胞分化为巨噬细胞并极化为M1巨噬细胞。
M1 巨噬细胞衍生的外泌体将 miR-326 递送至 HCC 细胞并影响 HCC 细胞中的 miR-326 表达
为了验证源自 M1 巨噬细胞的外泌体是否将 miR-326 转运至 HCC 细胞,将 HepG2 和 SMMC-7721 细胞与外泌体共培养。荧光显微镜下转染4h后,大量外泌体被HepG2和SMMC-7721细胞同化(图2a、b)。
<图片>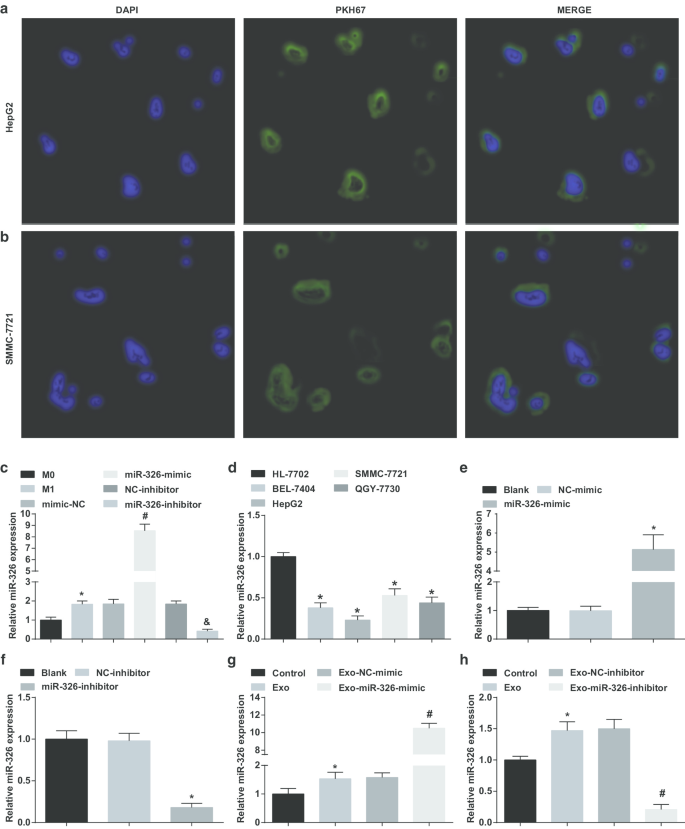
M1 巨噬细胞衍生的外泌体将 miR-326 递送至 HCC 细胞并影响 HCC 细胞中 miR-326 的表达。 一 HepG2 细胞摄取 M1 巨噬细胞衍生的外泌体。 b SMMC-7721 细胞对 M1 巨噬细胞衍生的外泌体的摄取。 c INF-γ和LPS诱导前后各组巨噬细胞外泌体miR-326表达比较。 d 通过 RT-qPCR 检测 HCC 细胞系(BEL-7404、HepG2、SMMC-7721、QGY-7703)和人正常肝细胞 HL-7702 细胞系中的 miR-326 表达。 e RT-qPCR 检测了 miR-326 模拟物对 HepG2 细胞中 miR-326 表达的影响。 f RT-qPCR检测了miR-326抑制剂对SMMC-7721细胞中miR-326表达的影响。 g RT-qPCR 检测了 miR-326 模拟转染的 M1 巨噬细胞衍生的外泌体对 HepG2 细胞中 miR-326 表达的影响。 h RT-qPCR 检测了 miR-326 抑制剂转染的 M1 巨噬细胞衍生的外泌体对 SMMC-7721 细胞中 miR-326 表达的影响。在面板 c 中,*P <0.05 与 M0 巨噬细胞相比;在面板 d 中,*P <0.05 与 HL-7702 细胞相比;在面板 e 中,*P <0.05 与 NC 模拟组相比;在面板 f 中,*P <0.05 与 NC 抑制剂组相比;在面板 g 中,*P <0.05 与对照组相比,#P <0.05 与 Exo-NC 模拟组相比;在面板 h 中,*P <0.05 与对照组相比,#P <0.05 与 Exo-NC 抑制剂组相比。测量数据被描述为平均值 ± 标准偏差(N =3),多组间比较采用单因素方差分析
随后,将 miR-326 模拟物/抑制剂转染到巨噬细胞中,并在转染前后检查其外泌体中 miR-326 的表达。结果表明,与来自 M0 巨噬细胞的外泌体相比,M1 巨噬细胞衍生的外泌体中 miR-326 表达升高。在用 miR-326 模拟物处理的 M1 巨噬细胞衍生的外泌体中观察到 miR-326 表达升高。 miR-326 抑制剂处理后 M1 巨噬细胞来源的外泌体中 MiR-326 表达降低(图 2c)。
然后,测试了 HCC 细胞系中的 miR-326 表达。如图所示,与 HL-7702 细胞相比,BEL-7404、HepG2、SMMC-7721 和 QGY-7703 细胞中 miR-326 表达降低,其中 HepG2 细胞表达最低,而 SMMC-7721 细胞表达最高(图2d)。
随后,将miR-326-mimic和miR-326-inhibitor分别转染到HepG2和SSMC-7721细胞中,并检测它们对miR-326表达的影响。 MiR-326模拟HepG2细胞中miR-326表达升高,而miR-326抑制剂降低SSMC-7721细胞中miR-326表达(图2e、f)。
接下来,将转染 miR-326-mimic 和 miR-326-inhibitor 的 M1 巨噬细胞的外泌体分别与 HepG2 和 SSMC-7721 细胞共培养。强调与外泌体共培养增加了 HCC 细胞中 miR-326 的表达,miR-326 模拟转染的 M1 巨噬细胞衍生的外泌体进一步提高了 HepG2 细胞中 miR-326 的表达,而 miR-326 抑制剂转染的 M1 巨噬细胞衍生的外泌体降低了 SSMC-7721 细胞中 miR-326 的表达(图 2g、h)。提示 M1 巨噬细胞来源的外泌体将 miR-326 递送至 HCC 细胞并影响 HCC 细胞中 miR-326 的表达。
M1 巨噬细胞衍生的外泌体 miR-326 降低 HCC 细胞中的细胞增殖和集落形成能力
在探索外泌体 miR-326 对 HCC 细胞增殖的影响时,进行了 MTT 和集落形成试验以检查 HCC 细胞增殖。这表明在 HepG2 细胞中,miR-326 的恢复损害了细胞增殖和集落形成能力(图 3a、e、f)。 M1 巨噬细胞衍生的外泌体阻碍 HepG2 细胞增殖并形成集落。 MiR-326模拟转染M1巨噬细胞来源的外泌体进一步损害细胞增殖和集落形成能力(图3c,i,j)。
<图片>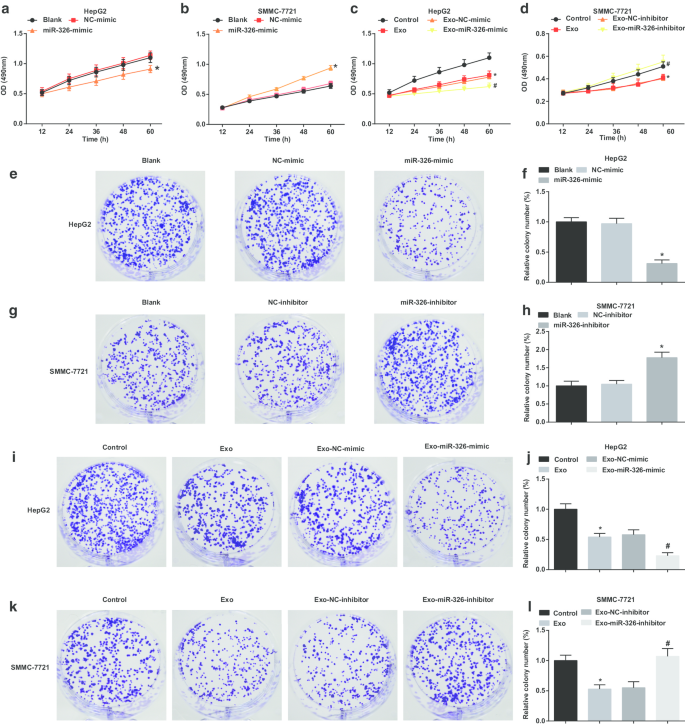
M1巨噬细胞外泌体miR-326抑制HCC细胞的细胞增殖和集落形成。 一 MTT法检测转染miR-326模拟物对HepG2细胞增殖的影响。 b MTT法检测转染miR-326抑制剂对SMMC-7721细胞增殖的影响。 c MTT法检测了与miR-326模拟物转染的M1巨噬细胞衍生的外泌体共培养对HepG2细胞增殖的影响。 d MTT法检测了与miR-326抑制剂转染的M1巨噬细胞来源的外泌体共培养对SMMC-7721细胞增殖的影响。 e 集落形成实验检测转染miR-326模拟物对HepG2细胞集落形成能力的影响。 f HepG2 细胞的集落数。 g 集落形成实验检测转染miR-326抑制剂对SMMC-7721细胞集落形成能力的影响。 h SMMC-7721 细胞的菌落数。 我 集落形成试验检测了与 miR-326 模拟物转染的 M1 巨噬细胞衍生的外泌体共培养对 HepG2 细胞集落形成能力的影响。 j 用外泌体处理的 HepG2 细胞的集落数。 k 集落形成实验检测了转染了 miR-326 抑制剂的 M1 巨噬细胞来源的外泌体共培养对 SMMC-7721 细胞集落形成能力的影响。 我 用外泌体处理的 SMMC-7721 细胞的菌落数。在面板 a 和 f 中,*P <0.05 与 NC 模拟组相比;在面板 b 和 h 中,*P <0.05 与 NC 抑制剂组相比;在面板 c 和 j 中,*P <0.05 与对照组相比,#P <0.05 与 Exo-NC 模拟组相比;在面板 d 和 l 中,*P <0.05 与对照组相比,#P <0.05 与 Exo-NC 抑制剂组相比。测量数据被描述为平均值 ± 标准偏差(N =3),多组间比较采用单因素方差分析
在 SMMC-7721 细胞中,miR-326 敲低增强了细胞增殖和集落形成能力(图 3b、g、h)。在用 M1 巨噬细胞衍生的外泌体处理的 SMMC-7721 细胞中,细胞增殖和集落形成能力降低。 MiR-326 抑制剂转染的 M1 巨噬细胞衍生的外泌体进一步促进细胞增殖和集落形成能力(图 3d,k,l)。提示M1巨噬细胞来源的外泌体miR-326阻碍HCC细胞增殖。
M1 巨噬细胞衍生的外泌体 miR-326 抑制 HCC 细胞的迁移和侵袭
然后,检查了外泌体 miR-326 对 HCC 细胞侵袭和迁移的影响。结果表明,在 HepG2 细胞中,miR-326 的恢复限制了侵袭和迁移(图 4a-c)。 M1 巨噬细胞衍生的外泌体干扰 HepG2 细胞入侵和迁移。当 HepG2 细胞与 miR-326 模拟转染的 M1 巨噬细胞衍生的外泌体共培养时,侵袭和迁移进一步降低(图 4g-i)。
<图片>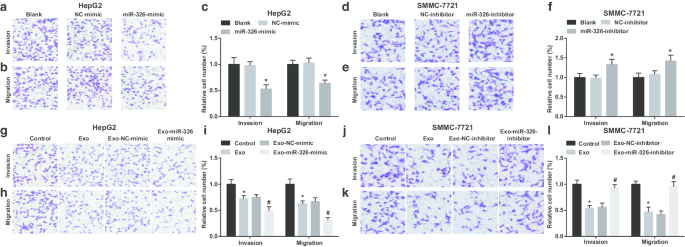
M1巨噬细胞外泌体miR-326抑制HCC细胞的迁移和侵袭。 一 Transwell实验检测转染miR-326模拟物对HepG2细胞侵袭的影响。 b Transwell 测定检测转染 miR-326 模拟 HepG2 细胞迁移的效果。 c HepG2 细胞侵袭和迁移的数量。 d Transwell实验检测转染miR-326抑制剂对SMMC-7721细胞侵袭的影响。 e Transwell实验检测转染miR-326抑制剂对SMMC-7721细胞迁移的影响。 f SMMC-7721细胞侵袭和迁移的次数。 g Transwell 实验检测了与 miR-326 模拟物转染的 M1 巨噬细胞衍生的外泌体共培养对 HepG2 细胞侵袭的影响。 h Transwell 试验检测了与 miR-326 模拟物转染的 M1 巨噬细胞衍生的外泌体共培养对 HepG2 细胞迁移的影响。 我 与 M1 巨噬细胞来源的外泌体共培养的 HepG2 细胞的侵袭和迁移次数。 j Transwell 实验检测了与 miR-326 抑制剂转染的 M1 巨噬细胞来源的外泌体共培养对 SMMC-7721 细胞侵袭的影响。 k Transwell 实验检测了与 miR-326 抑制剂转染的 M1 巨噬细胞来源的外泌体共培养对 SMMC-7721 细胞迁移的影响。 我 与 M1 巨噬细胞来源的外泌体共培养的 SMMC-7721 细胞的侵袭和迁移次数。在面板 c 中,*P <0.05 与 NC 模拟组相比;在面板 f 中,*P <0.05 与 NC 抑制剂组相比;在面板 i 中,*P <0.05 与对照组相比,#P <0.05 与 Exo-NC 模拟组相比;在面板 l 中,*P <0.05 与对照组相比,#P <0.05 与 Exo-NC 抑制剂组相比。测量数据被描述为平均值 ± 标准偏差(N =3),多组间比较采用单因素方差分析
MiR-326 敲低导致 SMMC-7721 细胞侵袭和迁移的增强(图 4d-f)。当用 M1 巨噬细胞衍生的外泌体处理时,SMMC-7721 细胞的侵袭和迁移减少。 However, SMMC-7721 cell invasion and migration were boosted upon co-culture with miR-326-inhibitor-transfected M1 macrophages-derived exosomes (Fig. 4j–l). It is implied that miR-326 derived from M1 macrophage exosomes impedes the invasion and migration of HCC cells.
M1 Macrophage-Derived Exosomal miR-326 Promotes Apoptosis of HCC Cells
When examining the effect of exosomal miR-326 on the cell cycle and apoptosis of HCC cells, PI single staining and Annexin V-FITC/PI double staining were applied. It was illustrated that miR-326 overexpression increased cells arrested at G0/G1 phase, reduced cells arrested at S and G2/M phases and raised apoptosis in HepG2 cells (Fig. 5a–d). Co-culturing with M1 macrophages-derived exosomes increased cells arrested at G0/G1 phase, reduced cells arrested at S and G2/M phases and raised cell apoptosis of HepG2 cells. Co-cultivation with exosomes from M1 macrophages transfected with miR-326-mimic further enhanced these effects (Fig. 5i–l).

M1 macrophage exosomal miR-326 promotes apoptosis of HCC cells. 一 , b Flow cytometry detected the effect of transfection of miR-326 mimic on HepG2 cell cycle; c , d flow cytometry detected the effect of transfection of miR-326 mimic on HepG2 cell apoptosis; e , f flow cytometry detected the effect of transfection of miR-326 inhibitor on SMMC-7721 cell cycle; g , h flow cytometry detected the effect of transfection of miR-326 inhibitor on SMMC-7721 cell apoptosis; 我 , j flow cytometry detected the effects of co-culture of with miR-326 mimic-transfected M1 macrophage-derived exosomes on HepG2 cell cycle; k , l flow cytometry detected the effect of co-culture with miR-326 mimic-transfected M1 macrophage-derived exosomes on HepG2 cell apoptosis; m , n flow cytometry detected the effect of co-culture with miR-326 inhibitor-transfected M1 macrophage-derived exosomes on SMMC-7721 cell cycle; o , p , flow cytometry detected the effect of co-culture with miR-326 inhibitor-transfected M1 macrophage-derived exosomes on SMMC-7721 cell apoptosis. In panel b and d, *P <0.05 versus the NC-mimic group; In panel f and h, *P <0.05 versus the NC-inhibitor group; In panel j and l, *P <0.05 versus the control group, #P <0.05 versus the Exo-NC-mimic group; In panel n and p, *P <0.05 versus the controlgroup, #P <0.05 versus the Exo-NC-inhibitor group. Measurement data were depicted as mean ± standard deviation (N = 3), comparisons among multiple groups were conducted by one-way analysis of variance
In SMMC-7721 cells, miR-326 down-regulation reduced cells arrested at G0/G1 phase, elevated cells arrested at S and G2/M phases, and declined cell apoptosis (Fig. 5e–h). Untransfected M1 macrophages-derived exosomes increased cells arrested at G0/G1 phase, reduced cells arrested at S and G2/M phases, and heightened cell apoptosis. MiR-326-inhibitor-transfected M1 macrophages-derived exosomes degraded cells arrested at G0/G1 phase, elevated cells arrested at S and G2/M phases, and decreased cell apoptosis (Fig. 5m–p). Briefly, it is summarized that miR-326 derived from M1 macrophage exosomes arrests cell cycle in G0/G1 phase and induces cell apoptosis in HCC.
M1 Macrophage-Derived Exosomal miR-326 Declines CD206 and NF-κB Expression in HCC Cells
Next, the potential mechanism of miR-326 derived from M1 macrophage exosomes in the biological progress of HCC cells was explored. NF-κB is the key link between inflammation and cancer. Many regulatory proteins and miRNAs could inhibit the excessively activated NF-κB signaling to suppress cancer [16]. Such a beneficial effect may include the polarization of M2 macrophages into M1 macrophages. CD206 and NF-κB expression in HepG2 and SMMC-7721 cells was tested by RT-qPCR. It was suggested that miR-326 restoration decreased CD206 and NF-κB expression in HepG2 cells, while miR-326 knockdown enhanced CD206 and NF-κB expression in SMMC-7721 cells (Fig. 6a, b). Moreover, co-culture with M1 macrophage exosomes significantly reduced CD206 and NF-κB expression in HepG2 cells, while co-culture with M1 macrophage exosomes-overexpressing miR-326 further decreased CD206 and NF-κB expression. Treated with untransfected M1 macrophages-derived exosomes, CD206 and NF-κB expression was decreased in SMMC-7721 cells. Co-cultured with miR-326-inhibitor-transfected M1 macrophages-derived exosomes, SMMC-7721 cells were featured by heightened CD206 and NF-κB expression (Fig. 6c, d). It was concluded that miR-326 from M1 macrophage exosomes played a tumor suppressor by inhibiting NF-κB in HCC cells.

M1 macrophage exosomal miR-326 declines CD206 and NF-κB expression in HCC cells. 一 RT-qPCR detected the effect of transfection of miR-326 mimic on the expression of CD206 and NF-κB in HepG2 cells; b RT-qPCR detected the effect of transfection of miR-326 inhibitor on the expression of CD206 and NF-κB in SMMC-7721 cells. c RT-qPCR detected the effect of co-culture with miR-326 mimic-transfected M1 macrophage-derived exosomes on the expression of CD206 and NF-κB in HepG2 cells; d RT-qPCR detected the effect of co-culture with miR-326 inhibitor-transfected M1 macrophage-derived exosomes on the expression of CD206 and NF-κB in SMMC-7721 cells. In panel a, *P <0.05 versus the NC-mimic group; In panel b, *P <0.05 versus the NC-inhibitor group; In panel c, *P <0.05 versus the control group, #P <0.05 versus the Exo-NC-mimic group; In panel d, *P <0.05 versus the control group, #P <0.05 versus the Exo-NC-inhibitor group. Measurement data were depicted as mean ± standard deviation (N = 3), comparisons among multiple groups were conducted by one-way analysis of variance
miR-326 from M1 Macrophage Exosomes Inhibits HCC Tumor Growth In Vivo
Finally, the in vivo results were validated through tumor xenografts. As displayed, miR-326 overexpression decreased volume and weight of tumors in HepG2 cells (Fig. 7a–c). In mice transplanted with HepG2 cells co-cultured with exosomes, the treatment with M1 macrophage exosomes significantly reduced the tumor volume and weight of HepG2 cells, while treatment with M1 macrophages-overexpressing miR-326 further reduced the tumor volume and weight of HepG2 cells (Fig. 7g–i).

M1 macrophage exosomal miR-326 reduces the volume and weight of HCC tumor in vivo. 一 –c The effect of transfection of miR-326 mimic on the tumor volume and tumor of nude mice xenografted with HepG2 cells. d –f The effect of transfection of miR-326 inhibitor on the tumor volume and tumors of nude mice xenografted with SMMC-7721 cells. g –我 The effect of co-culture with miR-326 mimic-transfected M1 macrophage-derived exosomes on the tumor volume and tumor of nude mice xenografted with HepG2 cells. j –l The effect of co-culture with miR-326 inhibitor-transfected M1 macrophage-derived exosomes on the tumor volume and tumor of nude mice xenografted with SMMC-7721 cells. In panel b and c, *P <0.05 versus the NC-mimic group; In panel e and f, *P <0.05 versus the NC-inhibitor group; In panel h and i, *P <0.05 versus the control group, #P <0.05 versus the Exo-NC-mimic group; In panel k and l, *P <0.05 versus the control group, #P <0.05 versus the Exo-NC-inhibitor group. Measurement data were depicted as mean ± standard deviation (n = 5), comparisons among multiple groups were conducted by one-way analysis of variance
In SMMC-7721 cells, miR-326 suppression increased volume and weight of tumors (Fig. 7d–f). Untransfected M1 macrophages-derived exosomes obstructed tumor growth in volume and weight of SMMC-7721 cells. Co-cultured with miR-326 inhibitor-transfected M1 macrophages-derived exosomes, SMMC-7721 cells were injected into mice and caused elevations in tumor volume and weight (Fig. 7j–l). Summarily, miR-326 derived from M1 macrophage exosomes depressed tumor growth of HCC in vivo.
Discussion
HCC is a common cancer that is characterized with high morbidity and mortality, difficult early diagnosis and treatment, poor prognosis and 5-year survival rate [17]. Recently, a study has highlighted that the lowly expressed lncRNA cox-2 declines the ability of M1 macrophages to suppress HCC cell growth, invasiveness, angiogenesis migration and promote apoptosis [18]. Hu et al. have discussed that miR-326 is obviously degraded in HCC tissues and cell lines, while down-regulated miR-326 is connected to the TNM stage, lymph node metastasis and differentiation of HCC patients [14]. It is customarily considered that HCC cells-derived exosomes can form a fertile environment to facilitate HCC cells growth, invasiveness and metastasis as well as development of drug resistance [19]. The current study was designed to explore the mechanism of exosomal miR-326 in regulating invasion and migration of HCC cells.
Our results indicated that miR-326 expression decreased in HCC cells but increased in exosomes. A recent study has pointed out that miR-326 expression is declined in HCC tissues [20]. Another study has presented miR-326 expression is notably reduced in HCC cell lines and tissues and its down-regulation predicts a poor prognosis in HCC [13]. It is reported that miR-326 acts a tumor-suppression role and is greatly depressed in HCC cells [21]. All these aforementioned evidences are in line with our findings. A study has purported that in comparison with the controls, miR-326 expression is raised in Tconv-derived exosomes which is observed in relapsing–remitting multiple sclerosis patients [11].
Other results emerge from our data that highly expressed exosomal miR-326 reduced cell proliferation, colony formation, migration and invasion as well as facilitated apoptosis of HCC cells in vitro and reduced the volume and weight of HCC tumor in vivo. It has been suggested previously that HCC cell growth can be suppressed via overexpression of miR-326, and HCC cell migration and invasion ability are markedly attenuated through elevating miR-326 [21]. It is reported that the up-regulated miR-326 expression suppresses HCC cell growth and invasiveness as well as stimulates cell apoptosis in vitro [14]. Besides that, a prior study has verified that overexpression of miR-326 declined tumor growth in vivo [13]. A study has revealed that ectopic expressed miR-326 markedly attenuates cell growth, and suppresses cellular migration and invasiveness in non-small cell lung cancer cell lines [22]. Moreover, it is found that miR-326 decreases profibrotic genes like MMP-9, implying its repressive function in cancer cell proliferation [23]. Also, it is presented that miR-326 represses Bcl-2 protein expression and elevates Bax expression so as to affect the apoptosis [24]. Similar to our findings, there are some miRNAs interacting with exosomes to play a role in HCC development. It is displayed that highly expressed exosomal miR-638 can repress the proliferation of HCC cells, involving the potential impact on carcinogenesis [25]. Another study also proves that HCC cells-derived exosomal miR-451a suppresses tumor angiogenesis via disrupting endothelial functions as apoptosis, tube formation, migration and permeability [26]. A prior research generally confirms that when treated with the overexpression of miR-744 exosomes, the proliferation of HCC cells is dramatically suppressed [27].
Conclusion
To briefly conclude, our study provides evidence that M1 macrophage-derived exosomal miR-326 suppresses proliferation, migration and invasion as well as advances apoptosis of HCC cells, supplying a new insight in a novel target therapy for HCC. Due to the limited sample size and limited known researches, the exact mechanism of miR-326 is not fully elucidated, and therefore, further large-scale studies are required to illustrate the underlying mechanism.
缩写
- HCC:
-
Hepatocellular carcinoma
- miRNAs:
-
MicroRNAs
- FBS:
-
胎牛血清
- M/CSF:
-
Macrophage colony-stimulating factor
- PBS:
-
磷酸盐缓冲盐水
- DMEM:
-
Dulbecco’s modified eagle medium
- PPAR:
-
Peroxisome proliferator-activated receptor
- TEM:
-
透射电子显微镜
- NTA:
-
Nanoparticle tracking analysis
- NC:
-
Negative control
- RT-qPCR:
-
Reverse transcription quantitative polymerase chain reaction
- BCA:
-
Bicinchoninic acid
- BSA:
-
牛血清白蛋白
- Exo:
-
Exosomes
- 方差分析:
-
单因素方差分析
纳米材料
- 通过介电超表面扩大带宽完成太赫兹偏振控制
- Au 纳米颗粒对 HT29 和 SPEV 细胞系影响的体外研究
- 斜角溅射 ITO 电极在 MAPbI3 钙钛矿太阳能电池结构中的影响
- 通过静电纺丝法制备用于直接甲醇燃料电池的新型复合二氧化钛碳纳米纤维阳极催化剂的制备和表征
- 通过聚乙二醇化脂质体改善蟾蜍灵的抗肿瘤功效和药代动力学
- 通过羟基调节碳纳米管的滑滚运动模式
- 评估金纳米颗粒抑制的细胞色素 P450 3A4 活性及其在人肝细胞癌细胞系 C3A 中的细胞毒性的分子机制
- 微小的稀土氟化物纳米粒子通过电极性相互作用激活肿瘤细胞生长
- 基于过氧化锌的可编程金属化单元中的开关失效机制
- 自噬抑制剂 (LY294002) 和 5-氟尿嘧啶 (5-FU) 基于组合的纳米脂质体,可增强对食管鳞状细胞癌的疗效
- 胃壁细胞和小肠杯状细胞分泌:一种新型细胞介导的体内金属纳米颗粒代谢途径通过中草药增强腹泻
- 通过金属氧化物纳米粒子调整细胞形态力学扰动