

SOX2 调控 lncRNA CCAT1/MicroRNA-185-3p/FOXP3 轴影响宫颈癌干细胞的增殖和自我更新
摘要
已经介绍了长链非编码 RNA (lncRNA) 在宫颈癌 (CC) 中的作用。我们旨在讨论性别决定区 Y-box 2 (SOX2)/lncRNA 结肠癌相关转录物-1 (CCAT1)/microRNA-185-3p (miR-185-3p)/forkhead box protein 3 (FOXP3) 的影响) 对 CC 干细胞的增殖和自我更新能力的影响。在 CC 组织和细胞中测试了 MiR-185-3p、SOX2、CCAT1 和 FOXP3 的表达。验证了 SOX2/CCAT1 表达与 CC 患者临床病理特征之间的关系。在 CD44 + 中进行了功能丧失和获得的研究 HeLa 细胞讨论生物功能和自我更新能力。最后,验证了 SOX2、CCAT1、FOXP3 和 miR-185-3p 之间的关系。 miR-185-3p 表达降低,而 SOX2、CCAT1 和 FOXP3 在 CC 组织和细胞中的表达增加。 SOX2 和 CCAT1 表达与肿瘤大小、淋巴结转移和国际妇产科联合会 CC 的分期有关。下调SOX2或CCAT1和上调miR-185-3p导致CD44 + 增殖、侵袭、迁移和细胞球数抑制以及凋亡加速 海拉细胞。 SOX2可以结合CCAT1影响miR-185-3p的表达,FOXP3被miR-185-3p靶向。
介绍
宫颈癌 (CC) 是女性第四大死亡原因,2018 年全球估计有 570,000 例病例和 311,000 例死亡[1]。这种复杂的疾病涉及多种因素,包括遗传效应和病毒感染[2]。随着人乳头瘤病毒联合检测和人乳头瘤病毒疫苗接种的发展,宫颈发育不良和癌症的早期诊断程序降低了 CC 的发病率、发病率和死亡率 [3]。对于早期CC患者,推荐手术,如保留生育能力手术、锥形活检、根治性宫颈切除术、盆腔淋巴结清扫、盆腔放疗和近距离放射治疗[4]。晚期CC患者由于转移或复发,预后仍较差[5]。因此,寻找新的、有效的预后标志物和治疗策略以改善CC的治疗仍是当务之急。
性别决定区Y-box 2(SOX2)是转录因子SOX家族的重要成员,主要表现在胚胎干细胞和成体干细胞中,也在肿瘤干细胞中表达[6]。研究表明,SOX2 通过刺猬信号通路调节 CC 的放射抗性 [7]。另一项研究表明,SOX2 对于维持 CC 细胞系中癌症干细胞的亚群至关重要 [8]。长链非编码 RNA (lncRNA) 是一类长度为 200 个核苷酸的 RNA 分子 [9]。 LncRNA 结肠癌相关转录物-1 (CCAT1) 位于人类染色体 8q24.21 上,被认为是导致结肠癌基因突变的“热点”[10]。一项研究报告称,CCAT1 可加速 CC 的细胞增殖和侵袭 [11]。根据 Jia 等人的说法,CCAT1 显着增强了 CC 细胞的增殖、迁移和侵袭 [12]。此外,另一项研究表明,CCAT1 通过减少 microRNA-185-3p (miR-185-3p) 破坏肠道屏障来增强炎症性肠病的恶性程度 [13]。 miRNA可以通过减少mRNA和抑制翻译来反向控制基因的表达[14]。之前的一项研究中的体外实验表明,miR-185-3p 可调节鼻咽癌的放射抗性 [15]。另一项研究表明,miR-185 在体内和体外参与了卵巢癌顺铂耐药性 [16]。叉头盒蛋白 3 (FOXP3) 是属于 FOX 蛋白家族的转录因子,首先在调节性 T (Treg) 细胞中发现,在 Treg 细胞的维持和过程中起着至关重要的作用 [17]。一项研究报告说 FOXP3 与 CC 的淋巴管生成有关 [18]。另一项研究表明,FOXP3 的水平与国际妇产科联合会分期(FIGO 分期)和 CC 的肿瘤大小密切相关 [19]。因此,本研究考察了SOX2/CCAT1/miR-185-3p/FOXP3轴对CC干细胞增殖和自我更新能力的影响。
材料和方法
道德批准和参与同意
涉及人类的实验是按照赫尔辛基宣言中表达的原则进行的。该研究获得了吉林大学第一医院机构审查委员会的批准。所有参与者均签署知情同意书。
学习科目
2016年12月至2018年12月,共采集CC患者39例CC组织及相应的邻近正常组织,液氮保存。纳入标准如下: (1)经宫颈管活检病理、宫颈涂片细胞学、宫颈碘试验、阴道窥器及颈椎切除术证实为CC。 (2) 患者术前 2 周未接受放疗和化疗。排除标准如下:(1)接受任何放疗或化疗的患者,(2)不同意样本采集的患者,(3)免疫系统疾病患者。
细胞选择和培养
CC 细胞系(SiHa、HeLa、CaSki、HCC94 和 C33A)和人宫颈上皮永生化细胞系 H8 购自上海生物叶生物技术有限公司(中国上海)。 CC 细胞 SiHa、HeLa 和 HCC94 在含有 10% 胎牛血清 (FBS) 的高糖 Dulbecco's Modified Eagle 培养基 (DMEM) 中培养,而 CaSki、C33A 和 H8 细胞在 Roswell Park Memorial Institute (RPMI)-1640 培养基中含有 10 % FBS(37℃和 5% CO2)。每2天分离细胞并传代一次。
CC 干细胞分选 (CD44 + 海拉细胞)
通过细胞悬浮培养将CC干细胞与CC细胞系HeLa分离。 CC 细胞 HeLa 通过无血清 Iscove 改良 Dulbecco 培养基 (IMDM) 在超低粘附培养皿中培养 21 天,每 3-5 天半定量更换培养基。将一些细胞悬浮成球体,获得 HeLa 球体形成细胞 (SFCs)。对球形细胞的性质进行了鉴定和分析。细胞用胰蛋白酶分离并调整至1 × 10 6 细胞/毫升。细胞加入CD44抗体并通过流式细胞术分选。 CD44阳性的HeLa细胞是HeLa肿瘤干细胞,而CD44阴性的HeLa细胞是HeLa非干细胞。 CC 干细胞在 DMEM/F12 中培养,并添加 20 ng/mL 碱性成纤维细胞生长因子 (bFGF)、20 ng/mL 表皮生长因子 (EGF) 和 B27。培养基含1%青霉素和链霉素[20]。
细胞治疗
CD44 + HeLa 细胞用 sh-SOX2、sh-SOX2 阴性对照 (NC)、sh-CCAT1、sh-CCAT1 NC、miR-185-3p 模拟物、模拟物 NC、sh-CCAT1 和 miR-185-3p 抑制剂以及sh-CCAT1 和抑制剂 NC。所有寡核苷酸序列均由 GenePharma(中国上海)提供。经胰蛋白酶分离后,将细胞接种于 3 × 10 6 的 6 孔板中 细胞/孔。当达到 60% 汇合时,将细胞更换为无血清培养基并孵育 1 小时。 Lipofectamine 2000转染试剂(Invitrogen,Carlsbad,California,USA)促进转染。
逆转录定量聚合酶链反应 (RT-qPCR)
组织和细胞中的 RNA 用 Trizol (Invitrogen) 提取。通过莫洛尼鼠白血病病毒 RTase 试剂盒(Invitrogen)将 RNA(1 μg)逆转为 cDNA。将 cDNA 添加到实时 PCR 系统中。引物由上海生工生物技术有限公司(中国上海)设计(表 1)。 U6 是 miR-185-3p 的上样对照,而 SOX2、CCAT1 和 FOXP3 的磷酸甘油醛脱氢酶 (GAPDH) 是上样对照。结果通过2 -ΔΔCt 进行分析 方法。
Western Blot Assay
提取细胞和组织中的总蛋白质。通过二辛可宁酸试剂盒(AmyJet Scientific,武汉,湖北,中国)测定蛋白质浓度。将蛋白质与上样缓冲液混合并煮沸 5 分钟,然后冰浴和离心。蛋白质用 10% 十二烷基硫酸钠-聚丙烯酰胺凝胶电泳处理并转移到膜上。膜用 5% 脱脂牛奶封闭 1 小时,用一抗 SOX2(1:1000,江苏瑞思坦有限公司,江苏,中国)、FOXP3(1:1000,Abcam Inc.,Cambridge,MA)探测, 美国)、GAPDH (1:1000, Cell Signaling Technology, Beverly, MA, USA) 并用辣根过氧化物酶标记的二抗重新检测,用保鲜膜覆盖并暴露。 Bio-rad Gel Doc EZ 成像仪(Bio-rad,加利福尼亚,美国)被用于开发。蛋白质图像通过ImageJ2x软件进行分析。
细胞计数试剂盒 (CCK)-8 分析
CCK-8 测定用试剂盒(Beyotime,上海,中国)实施。细胞 (1 × 10 4 ) 接种到 96 孔板中并孵育。培养 0、24、48 和 72 小时,将细胞加入 10 μL/孔 CCK-8 溶液并孵化 1 小时。使用 Multiskan Spectrum 全波长酶标仪在 450 nm 处测定光密度值。取6口井计算平均值。以时间为纵坐标,以相对细胞活力为纵坐标绘制细胞生长曲线。光密度值代表细胞增殖。
流式细胞术
细胞 (1 × 10 6 ) 以 1500 r/min 离心,用 200 μL 结合缓冲液悬浮,依次与 5 μL 碘化丙啶 (PI) 和 5 μL Annexin V-异硫氰酸荧光素 (FITC) 孵育,并附加 400 μL 缓冲液。流式细胞仪(BD Biosciences,NJ,USA)验证细胞凋亡率。
划痕测试
将转染的细胞分离并制备成细胞悬液。细胞悬液(1 × 10 6 细胞/mL) 接种在 6 孔板中并培养至 80-90% 汇合。 6 孔板上的横向和纵向划痕由 10 μL 无菌移液器吸头沿尺子均匀划伤。用完全培养基代替原培养基,继续培养细胞。 0 h和72 h显微镜下观察细胞迁移情况,并在同一部位拍照。
Transwell 检测
将转染的细胞用胰蛋白酶消化,以 1000 g 离心并用 DMEM 冲洗。将完全水合的 Transwell 室置于含有 10% FBS-DMEM(600 mL/孔)的 24 孔板中。细胞悬液(5 × 10 5 细胞/mL,200 μL)加入Transwell上室(涂有1:8 Matrigel 80 μL),而500 μL 20% FBS-DMEM加入下室。细胞连续培养 24 小时,用 500 μL 4% 多聚甲醛 (PFA) 固定并用 0.1% 结晶紫染色液染色。然后,用棉签擦去上室表面和边缘的细胞。随机选取5个视野,在Nikon Eclipse TE2000-S显微镜(Nikon,Japan)下对细胞进行计数。
球体形成分析
将细胞以 200 个细胞/孔接种在含有无血清悬浮培养基的低吸附 6 孔板中。 2 w后,在Nikon Eclipse TE2000-S显微镜(Nikon)下观察细胞球形成率,球形成率计算为平均球数/接种细胞数 × 100%。
染色质免疫沉淀 (ChIP) 检测
ChIP 检测使用 ChIP 试剂盒(美国纽约州北部)进行。 SOX2(1:1000,Re-stem Biotech,江苏,中国)和正常兔 IgG(12-370,Millipore,美国)抗体用于免疫沉淀交联的蛋白质-DNA 复合物。 CD44 + HeLa 细胞用 1% PFA 固定并孵育以产生 DNA-蛋白质交联。然后,通过超声波将 DNA 切割成 200-300 bp 的染色质片段。回收沉淀的染色质DNA并进行RT-qPCR分析。
RNA 下拉分析
将生物素标记的 miR-185-3p 野生型 (WT) 和突变型 (MUT) 质粒 (50 nM) 转染到 CD44 + HeLa 细胞。转染 48 小时后,用特定细胞裂解物(Ambion,Austin,Texas,USA)孵化细胞。细胞裂解物 (50 mL) 被分装。残留的裂解物用 M-280 链霉亲和素珠(Sigma,St. Louis,MO,USA)孵化,该珠用无 RNase 和酵母 tRNA(Sigma)预包被。然后,用冷裂解液清洗细胞两次,用低盐缓冲液清洗 3 次,用高盐缓冲液清洗一次。拮抗 miR-185-3p 探针被设置为 NC。 Trizol提取总RNA,RT-qPCR检测CCAT1水平。
双荧光素酶报告基因检测
通过 https://jaspar.genereg.net/ 预测 CCAT1 启动子区域上 SOX2 的潜在结合位点 E1 和 E2。合成含有SOX2和CCAT1结合位点E1的CCAT1启动子序列,形成CCAT1 3'UTR WT(E1-WT)和CCAT1 3'UTR MUT(E1-MUT)载体。将载体克隆到 pmirGLO (Beyotime) 中。之后,CCAT1-WT/pmirGLO 或 CCAT1-MUT/pmirGLO 与 sh-SOX2 或 sh-SOX2 NC 共转染 CD44 + HeLa细胞分别放置2 d后裂解。荧光素酶活性检测采用荧光素酶检测系统(Takara,大连,中国)。
利用生物信息学网站预测和分析 CCAT1 和 miR-185-3p 的结合位点。 CCAT1 和 miR-185-3p 的结合位点通过双荧光素酶报告基因检测进行验证。包含 miR-185-3p 结合位点的 CCAT1 3'UTR 组成。构建 CCAT1 3'UTR WT 和 CCAT1 3'UTR MUT 并与模拟物 NC 和 miR-185-3p 模拟物共转染至 CD44 + HeLa 细胞 2 天。然后,裂解细胞并通过荧光素酶检测系统(Takara)测试荧光素酶活性。应用相同方法验证miR-185-3p与FOXP3的靶向关系。
统计分析
所有数据均通过 SPSS 21.0 软件(IBM Corp. Armonk, NY, USA)进行评估。测量数据表示为平均值 ± 标准偏差。 t 检验适用于两组之间的差异和单因素方差分析 (ANOVA),然后是 Tukey 的多重比较检验以检测组间差异。分类变量通过 Fisher 精确检验进行评估。一个 p 小于0.05被认为是显着的。
结果
miR-185-3p 表达降低,而 CC 组织中 SOX2、CCAT1 和 FOXP3 表达增加,而 SOX2 和 CCAT1 表达与肿瘤大小、淋巴结有关转移 (LNM) 和 FIGO 分期
检测SOX2/CCAT1/miR-185-3p/FOXP3轴对CC干细胞增殖和自我更新能力的作用时,CC组织及邻近正常组织中miR-185-3p、SOX2、CCAT1和FOXP3的表达通过 RT-qPCR 和蛋白质印迹分析进行测试。结果表明(图1a-c)miR-185-3p表达降低,而CC组织中SOX2、CCAT1和FOXP3表达增加。
<图片>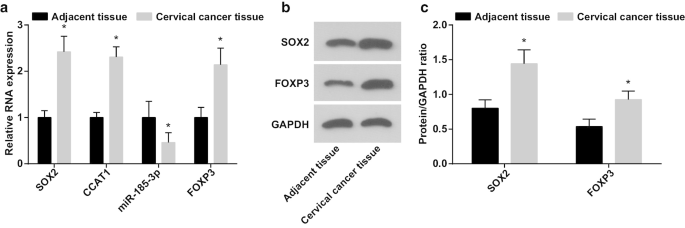
CC 组织中 miR-185-3p 表达降低,而 SOX2、CCAT1 和 FOXP3 表达增加。 一 CC和邻近正常组织中SOX2 mRNA、CCAT1、miR-185-3p和FOXP3 mRNA表达的比较。 b CC 和邻近正常组织中 SOX2 和 FOXP3 蛋白表达的蛋白条带。 c CC和邻近正常组织中SOX2和FOXP3蛋白表达的比较。 *p <0.05 与邻近的正常组织相比。计量资料以均值 ± 标准差表示,两组比较采用t检验
分析了 SOX2/CCAT1 表达与 CC 临床病理特征之间的关系(表 2)。 CC中SOX2和CCAT1的过表达与肿瘤大小、LNM和FIGO分期有关,说明SOX2和CCAT1在CC患者肿瘤较大、LNM和FIGO分期较大的患者中表达较高。
miR-185-3p 表达降低,而 CD44 中 SOX2、CCAT1 和 FOXP3 表达增加 + 海拉细胞
接下来,测试了人宫颈上皮永生化细胞 H8 和 CC 细胞系 SiHa、CasKi、HeLa、HCC94 和 C33A 中 miR-185-3p、SOX2、CCAT1 和 FOXP3 的表达。有人提出(图 2a-c)miR-185-3p 表达降低,而 SOX2、CCAT1 和 FOXP3 在 CC 细胞系中升高。其中HeLa细胞中SOX2/CCAT1/miR-185-3p/FOXP3与H8细胞表达差异最大;从而筛选出HeLa细胞并进行分选;细胞悬浮培养获得HeLa-SFCs,流式细胞仪检测分选前后细胞表面CD44的表达情况。结果强调,HeLa-SFCs中CD44的阳性率明显高于分选前,表明CC干细胞分选成功(图2d)。分选的干细胞命名为CD44 + 海拉细胞。然后,在HeLa和CD44 + 中测试miR-185-3p、SOX2、CCAT1和FOXP3的表达 HeLa 细胞(图 2e-g)。表明CD44 + 中SOX2、CCAT1和FOXP3表达上调,miR-185-3p下调 海拉细胞。
<图片>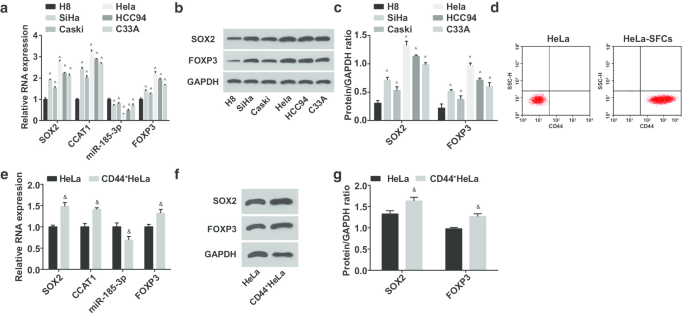
CD44 + 中miR-185-3p表达降低,而SOX2、CCAT1和FOXP3表达升高 海拉细胞。 一 H8 和 CC 细胞系中 SOX2 mRNA、CCAT1、miR-185-3p 和 FOXP3 mRNA 表达的比较。 b H8 和 CC 细胞系中 SOX2 和 FOXP3 蛋白表达的蛋白条带。 c H8 和 CC 细胞系中 SOX2 和 FOXP3 蛋白表达的比较。 d 流式细胞仪分选前后HeLa细胞CD44表达率检测。 e HeLa细胞与CD44 + SOX2 mRNA、CCAT1和miR-185-3p表达的比较 海拉细胞。 f HeLa细胞和CD44中SOX2蛋白表达的蛋白条带 + 海拉细胞。 g HeLa细胞与CD44细胞SOX2蛋白表达比较 + 海拉细胞。 ^p <0.05 对比 H8 细胞。 & p <0.05 对比 HeLa 细胞。测量数据以均值 ± 标准差表示,两组比较采用t检验,多组比较采用单向方差分析和Tukey's post hoc检验
下调 SOX2 和下调 CCAT1 降低增殖、迁移、侵袭和球形成率并增加 CD44 的细胞凋亡 + 海拉细胞
随后,SOX2 和 CCAT1 在 CD44 + 中沉默 HeLa 细胞探索其对 CC 干细胞增殖和自我更新的影响。通过CCK-8法、流式细胞术、划痕试验、Transwell试验和成球实验检测,CD44 + 的增殖、迁移、侵袭和成球率受到抑制,细胞凋亡 HeLa 细胞通过抑制 SOX2 和 CCAT1 得到促进(图 3a-i)。提示沉默SOX2或CCAT1可抑制CC干细胞的增殖和自我更新。
<图片>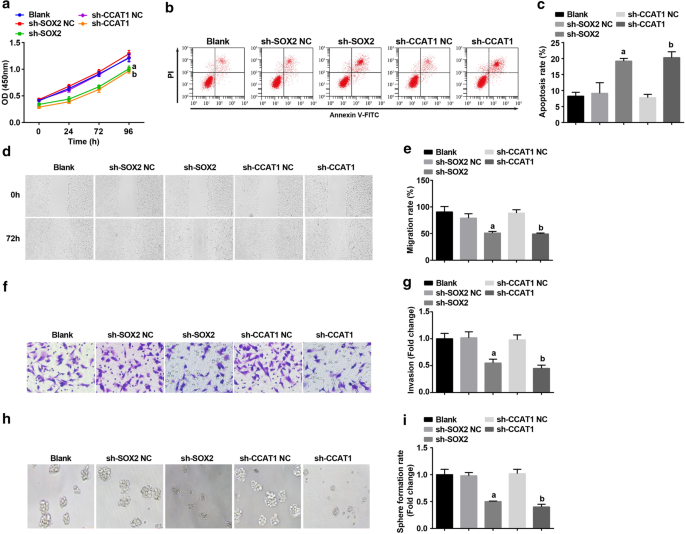
下调SOX2和下调CCAT1可降低CD44 + 的增殖、迁移、侵袭和球形成率,增加细胞凋亡 海拉细胞。 一 CCK-8 测定测试了用 sh-CCAT1 或 sh-SOX2 处理的细胞中的细胞生长曲线。 b 流式细胞术检测用 sh-CCAT1 或 sh-SOX2 处理的细胞中的细胞凋亡。 c sh-CCAT1 或 sh-SOX2 处理的细胞中细胞凋亡率的比较。 d 通过划痕测试测试用 sh-CCAT1 或 sh-SOX2 处理的细胞中的细胞迁移。 e sh-CCAT1 或 sh-SOX2 处理的细胞中细胞迁移的比较。 f Transwell实验检测sh-CCAT1或sh-SOX2处理后细胞的侵袭能力。 g sh-CCAT1或sh-SOX2处理的细胞侵袭能力的比较。 h 球体形成实验测试了用 sh-CCAT1 或 sh-SOX2 处理的细胞的自我更新能力。 我 sh-CCAT1 或 sh-SOX2 处理的细胞中球体形成率的比较。 一 p <0.05 vs. sh-SOX2 NC 组。 b p <0.05 vs. sh-CCAT1 NC 组。测量数据以均值 ± 标准差表示,多组间比较采用单向方差分析和Tukey事后检验
耗尽的 miR-185-3p 逆转 CCAT1 在 CD44 中下调的作用 + 海拉细胞
然后,我们检查了 miR-185-3p 是否参与了 CCAT1 调节 CC 干细胞增殖和自我更新的过程。 CD44 + HeLa 用 miR-185-3p 模拟物转染或与 sh-CCAT1 和 miR-185-3p 抑制剂共转染。结果表明,miR-185-3p的上调显着降低了CD44 + 的增殖、迁移、侵袭和球形成率,同时提高了细胞凋亡率。 海拉细胞。用miR-185-3p抑制剂处理的细胞可以逆转下调的CCAT1在CD44 + 增殖、迁移、侵袭、凋亡和细胞球形成中的作用 HeLa 细胞(图 4a-i)。
<图片>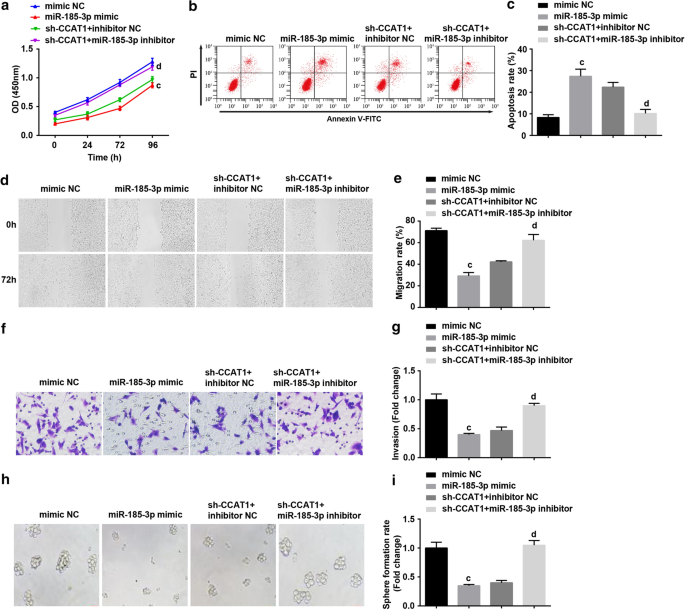
过表达miR-185-3p抑制CD44 + 的增殖、迁移、侵袭和球体形成率,增加细胞凋亡 海拉细胞。 一 CCK-8 测定测试了用 miR-185-3p 模拟物处理的细胞中的细胞生长曲线。 b 流式细胞术检测了用 miR-185-3p 模拟物处理的细胞中的细胞凋亡。 c 用 miR-185-3p 模拟物处理的细胞中细胞凋亡率的比较。 d 通过划痕测试测试用 miR-185-3p 模拟物处理的细胞中的细胞迁移。 e 用 miR-185-3p 模拟物处理的细胞中细胞迁移的比较。 f Transwell实验检测miR-185-3p模拟物处理细胞的侵袭能力。 g miR-185-3p模拟物处理的细胞侵袭能力的比较。 h 球体形成实验测试了用 miR-185-3p 模拟物处理的细胞的自我更新能力。 我 用 miR-185-3p 模拟物处理的细胞中球体形成率的比较。 p <0.05 vs. sh-SOX2 NC 组。 b p <0.05 vs. sh-CCAT1 NC 组。 c p <0.05 与模拟 NC 组相比。 d p <0.05 vs. sh-CCAT1 + 抑制剂NC组。测量数据以均值 ± 标准差表示,多组间比较采用单向方差分析和Tukey事后检验
低表达 SOX2 和低表达 CCAT1 在 CD44 中降低 FOXP3 表达并增加 miR-185-3p 表达 + 海拉细胞
之后,我们检测了 CD44 + 中 SOX2/CCAT1/miR-185-3p/FOXP3 的表达 转染 sh-SOX2、sh-CCAT1、miR-185-3p 模拟物并共转染 sh-CCAT1 和 miR-185-3p 抑制剂后的 HeLa 细胞。在用 sh-SOX2 处理的细胞中,SOX2、CCAT1 和 FOXP3 表达降低,而 miR-185-3p 表达升高。在用 sh-CCAT1 处理的细胞中,CCAT1 和 FOXP3 表达降低,miR-185-3p 表达增强。在引入 miR-185-3p 模拟物的细胞中,miR-185-3p 表达升高,FOXP3 表达降低。在依次转染sh-CCAT1和miR-185-3p抑制剂的细胞中,FOXP3表达升高,miR-185-3p表达降低(图5a-d)。
<图片>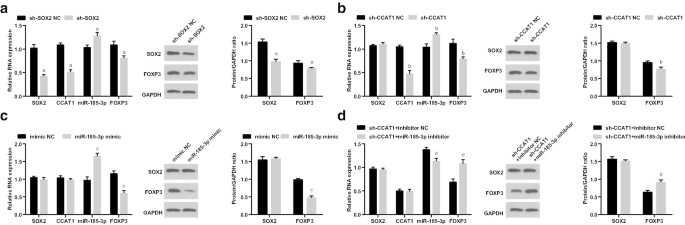
低表达的SOX2和低表达的CCAT1降低CD44 + 中FOXP3的表达并增加miR-185-3p的表达 海拉细胞。 一 SOX2/CCAT1/miR-185-3p/FOXP3 在 sh-SOX2 NC 和 sh-SOX2 组中的表达。 b SOX2/CCAT1/miR-185-3p/FOXP3 在 sh-CCAT1 NC 和 sh-CCAT1 组中的表达。 c SOX2/CCAT1/miR-185-3p/FOXP3 在模拟 NC 和 miR-185-3p 模拟组中的表达。 d SOX2/CCAT1/miR-185-3p/FOXP3在sh-CCAT1 + 抑制剂NC和sh-CCAT1 + miR-185-3p抑制剂组中的表达。 p <0.05 vs. sh-SOX2 NC 组。 b p <0.05 vs. sh-CCAT1 NC 组。 c p <0.05 与模拟 NC 组相比。 d p <0.05 vs. sh-CCAT1 + 抑制剂NC组。测量数据以均值 ± 标准差表示,多组间比较采用单向方差分析和Tukey事后检验
SOX2与影响miR-185-3p表达的CCAT1结合,FOXP3是miR-185-3p的靶基因
通过https://jaspar.genereg.net/网站预测和分析了CCAT1启动子区域转录因子的潜在结合位点,结果表明SOX2和CCAT1在CCAT1启动子区域具有潜在结合位点(图6a)。 ChIP-qPCR 报道(图 6b):与 IgG 组相比,SOX2 组在 E1 结合位点富集了更多的 CCAT1 启动子片段,这证明 SOX2 在 E1 位点与 CCAT1 启动子结合,而 SOX2 是参与 CCAT1 的调控。双荧光素酶报告基因检测显示(图6c):sh-SOX2和E1-WT共转染细胞中荧光素酶活性受到抑制,表明SOX2可以与CCAT1结合。
<图片>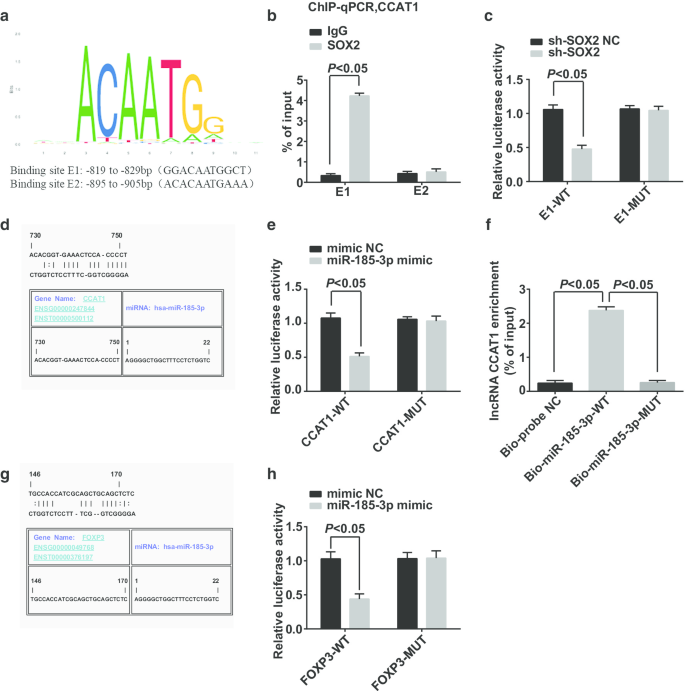
SOX2可以结合影响miR-185-3p表达的CCAT1,FOXP3是miR-185-3p的靶基因。 一 通过生物信息学位点预测 SOX2 和 CCAT1 启动子区域的结合位点。 b ChIP-qPCR 实验验证了 SOX2 与 CCAT1 的结合关系。 c SOX2 和 CCAT1 的结合位点通过双荧光素酶报告基因检测验证。 d 通过生物信息学位点预测 CCAT1 和 miR-185-3p 中的结合位点。 e 通过双荧光素酶报告基因检测验证 CCAT1 和 miR-185-3p 的结合。 f 通过RNA pull-down测定验证细胞中CCAT1和miR-185-3p之间的结合关系。 g 生物信息学网站对miR-185-3p与FOXP3靶向关系的预测。 h 双荧光素酶报告基因检测miR-185-3p与FOXP3靶向关系的鉴定计量资料以均值 ± 标准差表示,两组比较采用t检验
Jefferson 网站预测 CCAT1 可以与 miR-185-3p 结合(图 6d)。双荧光素酶报告基因测定报告(图 6e)在引入 miR-185-3p 模拟物和 CCAT1-WT 的细胞中荧光素酶活性降低,表明 miR-185-3p 可以与 CCAT1 结合。 RNA pull-down试验用于验证CCAT1是否可以与miR-185-3p结合。结果显示(图6f)Bio-miR-185-3p-WT处理的细胞中CCAT1的富集水平显着增加,而Bio-miR-185-3p-MUT处理的细胞中CCAT1的富集水平显示没有显着差异。该结果表明CCAT1可以吸附miR-185-3p,从而影响miR-185-3p的表达。
Jefferson 网站预测了 miR-185-3p 和 FOXP3 之间的靶标关系(图 6g)。双荧光素酶报告基因检测证实(图 6h)在 FOXP3-WT 和 miR-185-3p 模拟物共转染 CD44 + 后,细胞的相对荧光素酶活性显着降低 HeLa 细胞,而 FOXP3-MUT 与 miR-185-3p 模拟物共转染不影响细胞的相对荧光素酶活性,表明 miR-185-3p 靶向 FOXP3。
讨论
CC 是世界上第四常见的女性恶性肿瘤,其次是乳腺癌、结肠癌和肺癌 [3]。有报道称,SOX2阳性表达的CC细胞表现出癌症干细胞的特征[21]。一项研究报道,CCAT1 是与 CC 相关的重要致癌 lncRNA,并在 CC 细胞生长和侵袭中发挥促进作用 [11]。另一项研究表明,miR-185-3p 可以预测鼻咽癌的放射敏感性并调节癌细胞的生长和凋亡 [22]。已有报道,循环中针对 FOXP3 的自身抗体反映了宫颈病变的持续进展,可能是 CC 早期预后的潜在生物标志物 [23]。本研究旨在探讨SOX2/CCAT1/miR-185-3p/FOXP3轴如何影响CC干细胞的增殖和自我更新能力。
根据我们的发现,在与肿瘤大小、LNM 和晚期 FIGO 相关的 CC 组织和细胞中,SOX2 和 CCAT1 表达增加。功能证明,下调SOX2和CCAT1可降低CC干细胞的增殖、迁移、侵袭和球形细胞数量,并增加其凋亡。与我们的研究类似,SOX2 表达在 CC 中呈增加趋势 [24, 25]。 Moreover, SOX2 expression is also up-regulated in CC cells derived from cancer stem cells [26]. Overexpressed SOX2 was suggested to link with clinicopathological characteristics of patients with several types of cancer, not limited to CC. For example, it was suggested that up-regulated SOX2 shows in cervical squamous cell carcinoma patients staged in FIGO I-II [27]. Moreover, SOX2 expression is linked to LNM in oral squamous cell carcinoma [28]. When it comes to the molecular function of SOX2 for cancer progression, there is an observational work presenting that down-regulated SOX-2 suppresses cell migration and invasion of cervical squamous cell carcinoma [29]. Meanwhile, another research has offered a proof that up-regulated SOX2 enhances CC cell clonogenicity, proliferation and tumorigenicity in vitro and in vivo than control cells [30].
Concerning to the regulatory relation between SOX2 and CCAT1, an existed study has presented that silencing SOX2 markedly reduces CCAT1 mRNA level [31]. As to the role of CCAT1 in cancers, a study has showed that CCAT1 expression is markedly elevated in CC tissues versus in the adjacent normal tissues [11, 12]. Of note, CCAT1 overexpression in CC is positively related to the tumor size [12]. In terms of the role of CCAT1 in cancer cell activity, there is a research highlighting that overexpressed CCAT1 accelerates CC cell proliferation, colony formation and invasion [11]. Interestingly, a previous research has demonstrated that the cell viability, invasive and migratory abilities are declined via knocking down CCAT1 [12]. Anyway, the functional effect of SOX2 and CCAT1 in other cancers was similar to that in CC.
Afterward, our research revealed that CCAT1 could bind to miR-185-3p, the down-regulated CCAT1 in CC and overexpressing miR-185-3p suppressed the proliferation and self-renewal abilities of CC stem cells. It is reported that CCAT1 and miR-185-3p are negatively correlated [13]. Furthermore, a result reported that a reduction is seen in miR-185-3p expression in radioresistant nasopharyngeal carcinoma cases [22]. Regarding to the suppressive function of miR-185-3p in cancer cell aggressiveness, a study has revealed that up-regulation of miR-185-3p suppresses the invasive and metastatic properties of nasopharyngeal carcinoma cells [32]. Furthermore, Zou et al. have suggested that restored miR-185 represses breast cancer cell growth and invasion [33]. There is a article finding that up-regulation of miR-185 declines the proliferation, invasion and colony formation capacities of non-small cell lung cancer cells in vitro [34]. It is presented that in vitro cell proliferation, invasion and migration as well as in vivo tumor growth are suppressed via miR-185-overexpressing in non-small cell lung cancer cells [35]. From those studies, the anti-tumor role of miR-185-3p in the present study was consistent with previous researches.
To proceed, we unveiled that miR-185-3p targeted FOXP3, the overexpressed gene in CC to regulate CC stem cell activities. In fact, FOXP3, the regulator of SOX2 cancer stem-like cell marker in colon cancer [36], has been investigated in CC, showing an up-regulation in CC cells [19] [37]. It was evidenced that elevating FOXP3 promotes the formation of tumor spheres and stimulates the stemness of non-small cell lung cancer cells [38].
Conclusion
Collectively, we explored for the first time that SOX2 transcription could activate CCAT1, thereby inhibiting miR-185-3p and regulating FOXP3 to promote the proliferation and self-renewal of CC stem cells, which is a potential avenue to treat CC. Additionally, however, limitations in this present study still exist in the relatively small trial size in the designed experiment. Thus, clinical researches might be further carried out to detect the efficacy for the treatment of CC.
数据和材料的可用性
不适用。
纳米材料
- 用于灵敏快速检测卵巢癌细胞的柔性石墨烯生物传感器的演示
- 用于癌症治疗的纳米粒子:当前的进展和挑战
- 用于癌症应用的基于细胞的药物递送
- 纳米技术灭活癌症干细胞
- TiO2 中金纳米粒子分布对染料敏化太阳能电池光学和电学特性的影响
- 氧化石墨烯杂交 nHAC/PLGA 支架促进 MC3T3-E1 细胞的增殖
- 关于双层/三层宽带 SiO2 抗反射膜横截面形貌的 TEM 和 STEM 研究
- 通过多元醇介导工艺制备和表征 ZnO 纳米夹
- 囊泡和细胞的调频波介电泳:以交叉频率周期性 U 形转弯
- 嵌入TiO2致密层的不同尺寸和浓度的Ag纳米颗粒对钙钛矿太阳能电池转换效率的影响
- 石胆酸修饰的金纳米粒子对肝癌细胞的凋亡作用
- 影响冲压过程成功和安全的因素